

خلاصه
هدف اصلی زیرساخت سبز (GI) ارائه شبکههای کاربردی از زیستگاهها و اکوسیستمها برای حفظ تنوع زیستی درازمدت، در عین حال بهینهسازی عملکردها و خدمات چشمانداز و اکوسیستم برای رفع نیازهای انسان است. به طور سنتی، مطالعات اتصال توسط بومشناسی حرکت با ویژگیهای گونهای خاص از نوع و زمان حرکت (به عنوان مثال، پراکندگی، جفتگیری، جفتگیری) و فواصل حرکت اطلاعرسانی میشود، در حالی که دادههای محیطی فضایی به ترسیم مسیرهای حرکت در سراسر مناظر کمک میکنند. تا به امروز، طیف وسیعی از روشها و رویکردها موجود است که (الف) در هر ارگانیسم و نوع حرکتی مستقل از مقیاسهای زمانی و مکانی مرتبط هستند، (ب) به عنوان نرمافزار رایگان مستقل یا پیادهسازی سفارشی GIS آماده استفاده هستند. و (ج) خروجی های بصری جذابی تولید کند که ارتباط با مدیران زمین را تسهیل می کند. با این حال، برای افزایش استحکام ارزیابیهای اتصال و اطمینان از اینکه روندهای فعلی در مدلسازی اتصال با پتانسیل کامل خود به GI کمک میکنند، مخرجهای مشترکی لازم است که بر اساس آنها برنامهریزی و استراتژیهای طراحی پایهگذاری شود. به همین ترتیب، ارزیابیهای اتصال قابل مقایسه و قابل تکرار برای اعمال نتایج این ابزارهای علمی برای طرحها و اجرای GI چند منظوره مورد نیاز است. در این مقاله، استفاده و محدودیتهای روشهای ارتباطی پیشرفته در کمک به پیادهسازی GI را مورد بحث قرار میدهیم. مخرج مشترکی که بر اساس آنها برنامه ریزی و استراتژی های طراحی مورد نیاز است. به همین ترتیب، ارزیابیهای اتصال قابل مقایسه و قابل تکرار برای اعمال نتایج این ابزارهای علمی برای طرحها و اجرای GI چند منظوره مورد نیاز است. در این مقاله، استفاده و محدودیتهای روشهای ارتباطی پیشرفته در کمک به پیادهسازی GI را مورد بحث قرار میدهیم. مخرج مشترکی که بر اساس آنها برنامه ریزی و استراتژی های طراحی مورد نیاز است. به همین ترتیب، ارزیابیهای اتصال قابل مقایسه و قابل تکرار برای اعمال نتایج این ابزارهای علمی برای طرحها و اجرای GI چند منظوره مورد نیاز است. در این مقاله، استفاده و محدودیتهای روشهای ارتباطی پیشرفته در کمک به پیادهسازی GI را مورد بحث قرار میدهیم.
کلید واژه ها:
شبکه زیست محیطی ; راهرو ; مانع ; اکولوژی جنبش ; سطح هزینه ؛ ارزیابی مدل
1. معرفی
محیطها توسط انسانها در سطح جهانی و با سرعتهای بیسابقهای در دوران آنتروپوسن تغییر شکل داده شده است [ 1 ، 2 ، 3 ]. اکوسیستم ها و موجودات به طور فزاینده ای تحت استرس از دست دادن کیفیت، اندازه و اتصال زیستگاه ها قرار می گیرند. کاهش سطح زیستگاه به عنوان محدودکننده تنوع زیستی گونهها [ 4 ، 5 ، 6 ] و خدمات اکوسیستم [ 7 ، 8 ] شناخته شده است، در حالی که تکه تکه شدن زیستگاه موفقیت حرکت را محدود میکند [ 9 ، 10 ]، و بنابراین جریان ژن را کاهش میدهد ([ 11 ]، اما به لقمان مراجعه کنید). و همکاران [ 12]). مناطق حفاظت شده به تنهایی تنها راه حل های محدودی برای چنین خطراتی ارائه می دهند، زیرا از یک سو، از تغییرات جهانی مستثنی نیستند و از سوی دیگر، مناطق حفاظت شده به تنهایی ممکن است منطقه زیستگاه کافی را حفظ نکنند یا شبکه های متصل کافی برای حفظ اهداف تنوع زیستی را تشکیل ندهند. بلند مدت [ 13 ، 14 ]. بنابراین، رویکردهای یکپارچه برای متعادل کردن تضادهای رو به رشد بین اکوسیستمهای طبیعی و اکوسیستمهای انسانی که توسط چارچوب مفهومی زیرساخت سبز (GI) در نظر گرفته شده است، جهتهای امیدوارکنندهای را ارائه میدهد [ 15 ، 16 ] ( شکل 1 ). در حالی که GI یک اصطلاح کاملاً تعریف شده نیست و جنبه های زیادی دارد [ 15 ، 16]، عمدتاً بر این اصل استوار است که “حفاظت و تقویت طبیعت و فرآیندهای طبیعی […] آگاهانه در برنامه ریزی فضایی و توسعه سرزمینی ادغام می شوند” [ 17 ، 18 ].
بر این اساس، GI به عنوان “شبکه برنامه ریزی شده استراتژیک از مناطق طبیعی و نیمه طبیعی با سایر ویژگی های محیطی طراحی شده و مدیریت شده برای ارائه طیف گسترده ای از خدمات اکوسیستم” در محیط های روستایی و شهری تعریف می شود [ 17 ]. تاکید زیادی بر خدمات اکوسیستم با دامنه ارائه طیف وسیعی از مزایای زیست محیطی، از جمله نگهداری و بهبود عملکردهای زیست محیطی داده شده است. عناصر GI در زمینه راه حل های مبتنی بر طبیعت مهم هستند، به عنوان مثال برای ارائه خدمات اکوسیستمی که آسیب پذیری سیستم ها را کاهش می دهد [ 19 ، 20 ]، یا برنامه ریزی برای سازگاری با آب و هوای شدید و آب و هوای متغیر [ 21 ، 22 ، 23 ].
در زمینه اتصال اکولوژیکی، GI شبکههای کاربردی از زیستگاههای پراکنده فضایی را برای حفظ تنوع زیستی و بهینهسازی عملکردها و خدمات اکولوژیکی اکوسیستمها [ 24 ] ارائه میکند ( شکل 1 ). بنابراین، GIهای موفق چند وجهی و فضایی هستند تا تعاملات اکولوژیکی در مناظر تحت تسلط انسان [ 15 ] را با در نظر گرفتن ویژگیهای زنده ماندن و تاریخ حیات گونهها مانند پتانسیلهای حرکتی آنها تسهیل کنند. GI ذاتاً برای درک پیکربندی و دینامیک منظر به داده های مکانی-زمانی متکی است (برای مثال Bartesaghi-Koc و همکاران [ 25 ] الگوهای مکانی-زمانی در زیرساخت سبز را به عنوان محرک تغییر دمای سطح زمین در نظر گرفتند و Liu, XL, et al. [26 ] از داده های مکانی-زمانی برای ارزیابی فرآیندهای شهرنشینی استفاده کرد. از آنجایی که گونهها از نظر نیازها و دامنه حرکت متفاوت هستند، راهروها برای برخی گونهها ممکن است مانعی برای برخی دیگر باشد. در حالی که اتصال ساختاری به عناصر منظری اشاره دارد که به طور بالقوه زیستگاه ها را بر اساس پیکربندی و ترکیبات یک منظره به هم متصل می کنند، اتصال عملکردی به فرآیندهای زیستی مهاجرت، پراکندگی و جریان ژن اشاره دارد، به عنوان مثال، توانایی تبادل افراد یا ژن ها در سراسر یک منظر [ 27 ] . 28]. بنابراین، اتصال عملکردی، اتصال مؤثر از دیدگاه گونه است. بنابراین، تحقیق در مورد نقش ساختار چشمانداز در ایجاد اتصال عملکردی تحت فشارهای متغیر مانند کاربری زمین و زمینههای اجتماعی-اقتصادی و همچنین تغییرات آب و هوایی مورد نیاز است [ 29 ]. به این ترتیب، مهم است که GI به طور فعال در برنامه ریزی فضایی ادغام شود تا از تداوم طولانی مدت اتصال عملکردی اطمینان حاصل شود [ 17 ]. علاوه بر این، رویکردهای جغرافیایی را می توان در ابزارهای پشتیبانی تصمیم برای GI و برنامه ریزی فضایی (همانطور که توسط Manna و همکاران [ 30 ] در مناظر زیتون، و توسط Stessens و همکاران [ 31 ] برای خدمات اکوسیستم شهری انجام شد) پیاده سازی کرد.
در این مقاله، فرصتها و چالشهای ارزیابیهای ارتباطی پیشرفته برای GI را از دیدگاه دادهها، مفاهیم و پیشرفتهای روششناختی شناسایی میکنیم. ما این کار را ابتدا با توضیح روشهایی که ارزیابیهای ارتباطی حرکت را مشخص میکنند، انجام میدهیم. در مرحله بعد، روندهای کمی سازی اتصال را شناسایی و تشریح می کنیم. از آنجا ما در مورد استراتژیهای برنامهریزی و طراحی برای GI بحث میکنیم، و پیشنهاد میکنیم که چگونه ارزیابیهای اتصال میتوانند کمک کنند. این در نهایت منجر به طرح راههایی برای GI پایدار برای اطمینان از تداوم اتصال طولانیمدت در مقیاس چشمانداز برای حفظ و ارتقای تنوع زیستی میشود.
2. مشخص کردن اتصال عملکردی
2.1. جنبش
حرکت ارگانیسم ها فرض اساسی در ارزیابی اتصال است. حرکت در تمام جانوران مشترک است و بر طیف وسیعی از فرآیندهای بیولوژیکی تأثیر میگذارد، با پیامدهای بسیاری از ارگانیسمهای فردی گرفته تا جمعیتها یا کل اکوسیستمها. هر حرکتی نتیجه مجموعه ای از علل پیچیده است که ریشه در بین گونه ها دارد (به عنوان مثال، رفتار اجتماعی، ویلیامز و همکاران، [ 32 ]) یا در زمینه گونه-محیط، یا هر دو. حرکت را می توان در تمام مقیاس های زمانی مشاهده کرد که بر یک یا چند نسل یا مقیاس های فضایی شامل افراد منفرد، طیف وسیعی از جمعیت ها یا توزیع مکانی یک گونه تأثیر می گذارد ( شکل 2)آ). به عنوان مثال، حرکت درون جمعیتی و حرکت کوتاهمدت ممکن است به تغذیه یا علامتگذاری قلمروها اشاره داشته باشد، در حالی که حرکت بین جمعیتی در مقیاسهای زمانی گسترده و زمانی طولانی، در نهایت تبادل ژنتیکی را تعیین میکند ( شکل 2 A). واضح است که تعریف انواع مختلف حرکت یک چالش باقی می ماند [ 33 ]، و حرکت ممکن است همیشه به طور منظم در یک نوع حرکت دسته بندی نشود (اما به LaPoint و همکاران [ 34 ] و McClure و همکاران، [ 35 مراجعه کنید)]). بنابراین برای به دست آوردن استنتاج اکولوژیکی صحیح از حرکت، تحلیل استفاده و محدودیتهای مفروضات و مفاهیم اساسی که به عنوان جایگزینی برای حرکت تعریف نشده استفاده میشوند، کلیدی است. در حالی که نقش محیط در شکلدهی به حرکت یکی از عمیقترین تحقیقات انجام شده است، به عنوان مثال، [ 36 ، 37 ، 38 ]، شواهد قانعکنندهای وجود دارد که اطلاعات بیولوژیکی اضافی مانند تراکم جمعیت، ظرفیت حمل محلی، یا فرآیندهای درونزا مانند گونهها وجود دارد. ویژگی های خاص تاریخ زندگی (به عنوان مثال، سیستم های جفت گیری، زمان تولید) در مسیرهای حرکتی مهم هستند [ 39 ، 40 ، 41 ]]. بنابراین مطالعات اتصال (به طور ضمنی) سوالاتی مانند چرا حرکت؟ چگونه، کجا و چه زمانی حرکت کنیم؟ محرک های احتمالی حرکت چیست؟ چارچوبی توسط ناتان و همکاران. [ 42 ] حول خود حرکت متمرکز است و به سه مؤلفه اساسی مربوط به فرد (حالت داخلی، حرکت و ظرفیت ناوبری) و مؤلفه چهارم اشاره دارد که به عوامل محیطی تعیین کننده مسیرهای حرکت اشاره دارد ( شکل 2 B). وضعیت درونی ارگانیسم انگیزه حرکت را مشخص می کند (به عنوان مثال، رفتار [ 43])، در حالی که ظرفیت حرکت، ظرفیت کلی ارگانیسم برای حرکت را از منظر بیومکانیکی منعکس می کند، و ظرفیت های ناوبری ارگانیسم (به عنوان مثال، توانایی های شناختی و حسی) نشان می دهد که کجا باید حرکت کرد. از دیدگاه یک حیوان، اهمیت نسبی این چهار جزء که حرکت را تعیین میکنند، بین موجودات زنده، اما همچنین در طول عمر موجودات متفاوت است، مانند تغییرات در تناسب اندام، یا محیط، شکارچیان یا فراوانی رقبا. بنابراین، “حرکت” فرآیند پیچیده ای است که پارامترسازی آن در رویکردهای اتصال، عوامل برون زا و درون زا را در نظر می گیرد که بسته به گونه و نوع حرکت در نظر گرفته شده در تحلیل ها، الگوهای حرکت را به درجات مختلفی هدایت می کند.
2.2. مقیاس داده، فضا و زمان برای حرکت
ارزیابی های اتصال به منابع داده های مختلف برای ارزیابی حرکت تکیه می کنند ( جدول 1 ). در تجزیه و تحلیل اکتشافی خود، سه هندسه داده را دسته بندی کردیم: نقطه، چندضلعی و چند خط، که ممکن است بسته به تحلیل، به داده های شطرنجی نیز تبدیل شوند. نقاط یا چندضلعی حضور گونهها و زیستگاه بهدستآمده از برنامههای پایش یا بررسیها شاید سادهترین نوع دادههای مورد استفاده در مطالعات اتصال باشند. داده های حضور گونه ها (نقطه ای) به طور سنتی هسته اصلی ارزیابی توزیع زیستگاه است.
بدیهی است که زیستگاه مناسب (چه به صورت نقشههای پتانسیل پیوسته (راستر) یا چند ضلعی یا چند خطی با تناسب مشابه) ممکن است نقش کلیدی در الگوهای حرکت گونهها ایفا کند، زیرا ممکن است مهاجران را جذب کند و در عین حال به عنوان منبع مهاجرت عمل کند. زیستگاه پرجمعیت می شود [ 44 ]. در حالی که Fattebert و همکاران. [ 45 ] نشان می دهد که مناسب بودن زیستگاه ممکن است یک نماینده مناسب برای ترجیح حرکت در طول پراکندگی برای برخی از گونه ها باشد، مشکلات زیادی برای استفاده از پروکسی های حرکت وجود دارد که به حرکت واقعی اندازه گیری شده متکی نیستند [ 46 ]]. به عنوان مثال، تناسب زیستگاه معمولاً فقط یک یا دو بعد ساکن از اکولوژی یک ارگانیسم (مثلاً پرورش، جویدن) را در بر می گیرد و بنابراین ممکن است فرآیندهای رفتاری یا مقیاس وسیع تری را که حرکت را هدایت می کند، منعکس نکند [ 47 ، 48 ]. همچنین، افراد پراکنده ممکن است نیازی به زیستگاهی نداشته باشند که از ساکنان دائمی پشتیبانی کند [ 49 ]. بنابراین، این سوال باقی می ماند که آیا زیستگاه مناسب (یا زیستگاه نامناسب [ 50 ، 51 ]) ممکن است تعیین کننده حرکت باشد یا خیر. بنابراین، برای اطمینان از اثربخشی و کاربرد نتایج تجزیه و تحلیل، پیش نیاز است که نتایج حرکت ناشی از دادههای حضور ساده یا زیستگاه مناسب مدلسازی شده را در یک زمینه رفتاری قرار دهیم [ 34 ,52 ]. نظرات کارشناسان منبع داده دیگری است، به طور کلی به شکل چند ضلعی، که بر اساس آن ارزیابی حرکت انجام می شود ( جدول 1 ). کارشناسان مشاهدات محلی را با جزئیات و اغلب بلندمدت اما نسبتاً ذهنی در مورد راهروهای حیات وحش یا زیستگاه جمع آوری می کنند. همچنین، دانش محلی اغلب برای ارتقاء مقیاس به مقیاس چشم انداز دشوار است. در حالی که برخی از مطالعات اتصال منحصراً حول نظرات متخصصان [ 53 ] تنظیم شدهاند، برخی دیگر نظرات متخصص را به عنوان اطلاعات تکمیلی مهم در مورد حرکت اعمال میکنند [ 11 ، 54 ]، با این حال برخی دیگر ادعا میکنند که رویکردهای آماری به وضوح از دانش متخصص بهتر عمل میکنند [ 55 ، 56 ]. همچنین از نظرات کارشناسان برای تعیین مقاومت انواع پوشش زمین استفاده می شود.57 ] یا برای تعریف روابط و احتمالات انتقال بین زیستگاه ها [ 58 ]. دانش محلی بلند مدت ارائه شده توسط کارشناسان منبعی غیر قابل جایگزین برای بینش در مورد سؤالات مرتبط است که به حل چالش های حفاظتی در دنیای واقعی کمک می کند [ 59 ، 60 ]، و همچنین بازخورد مهم در مورد کاربرد و اعتبار ارزیابی های اتصال در مقیاس بزرگ به صورت محلی [ 59، 60]. 11 ].
نوع دیگری از داده های حرکتی به داده های چند خطی اشاره دارد که توسط ردیابی GPS یا تله متری ارزیابی می شود. به ویژه ردیابی GPS اجازه می دهد تا اطلاعات مکانی و زمانی دقیقی در مورد استفاده افراد از منظره به شکل چندخط ارائه شود و ممکن است اطلاعاتی در مورد رفتار یک حیوان اضافه کند ( جدول 1 ). در عین حال، داده های ردیابی تله متری یا GPS در مورد تأثیرات اتصال مشاهده شده برای تداوم جمعیت اطلاعی ندارند. دومی توسط داده های ژنتیکی، به عنوان چند خطوط استنباط شده و خوشه بندی فضایی تضمین می شود ( جدول 1) که امکان تعیین مستقیم اینکه آیا و تا چه اندازه جریان ژن تحقق یافته است و چگونه حرکت بر تمایز ژنتیکی تأثیر گذاشته و به تنوع ژنتیکی کمک کرده است را می دهد. جریان ژن را می توان در اعماق زمانی مختلف تجزیه و تحلیل کرد. مهاجران نسل اول ممکن است حرکت کوتاه مدت را در طول فصل اخیر از جمله جهت مبادله نشان دهند [ 62 ، 63 ]، در حالی که آزمایش های تخصیص ژنتیکی نشان دهنده حرکت در گذشته نزدیک است [ 64 ، 65 ]. تمایز ژنتیکی fst ، که یک معیار ژنتیکی نشان دهنده فرآیندهای تاریخی در نظر گرفته می شود (به عنوان مثال، بیش از 20 نسل)، می تواند برای شناسایی مسیرهای حرکت در اعماق زمانی بیشتر استفاده شود [ 66 ]]. هیچ عمق زمانی، اما خوشه بندی صریح فضایی شباهت ژنتیکی را می توان شناسایی کرد [ 67 ]. برای ارزیابی اتصال، اطلاعات به دست آمده از چنین الگوریتم های خوشه بندی نشان می دهد که چگونه ترکیب ژنتیکی ممکن است در یک چشم انداز متفاوت باشد [ 68 ]. علاوه بر این، بسیاری از مطالعات ژنتیکی معمولاً معیارهای دقیق تری از جمعیت مانند تنوع ژنتیکی یا اندازه جمعیت را در نظر می گیرند [ 63 ، 69 ، 70 ].
ما نتیجه میگیریم که بین دادههای حرکت در رابطه با اطلاعات مربوط به فرآیندهای حرکت (تولید مجدد در مقابل جزئیات مسیرهای حرکت تحققیافته در سراسر چشمانداز در مقابل اطلاعات در مورد توزیع زیستگاه) معاوضههایی وجود دارد. در حالی که دادههای ژنتیکی مشخص میکنند که آیا حرکت بر تولید مثل یا تنوع ژنتیکی تأثیر دارد یا خیر، و اجازه تمایز زمانی حرکت را میدهد، نوع داده مستقیماً در مورد مسیرهای حرکتی تحقق یافته (پلی خطوط) در سراسر مناظر اطلاعاتی نمیدهد. در مقابل، ردیابی GPS یا تله متری بینش هایی را در مورد مسیرهای حرکتی دقیق افراد در سراسر چشم انداز با وضوح مکانی و زمانی بالا ارائه می دهد. با این حال، دادهها اغلب نشاندهنده الگوهای حرکتی جزئی تنها چند نفر بین جمعیتها یا افراد بدون اطلاعات در مورد موفقیت باروری و جریان ژن هستند. مشاهدات حاصل از نظارت یا بررسیها صرفاً نشان میدهد که گونه در زمان بررسی (نقاط) حضور داشته است، اما میتواند نمونه بزرگتری از جمعیت را در یک منطقه معین نشان دهد و اغلب جمعآوری میشود. با این حال، این دادهها مستقیماً بر مسیرها و جهتهای حرکت یا اینکه آیا این حرکت به موفقیت باروری منجر میشود منعکس نمیشود و احتمالاً ضعیفترین نوع داده مورد استفاده در ارزیابیهای اتصال است.
به عنوان مثال، فری و همکاران. [ 71 ] نشان داد که دادههای پایش تنها حرکات ناچیزی را در وزغهای ناترجک شناسایی میکند، در حالی که ترکیبی از دادههای ردیابی، ژنتیکی و نظارت به طور قابلتوجهی نرخ تشخیص حرکت در سراسر چشمانداز را افزایش میدهد. این نشان میدهد که نوع دادههای حرکتی و هندسه مورد استفاده برای ارزیابی اتصال، نوع حرکتی را که میتوان در تحلیل تشخیص داد، دیکته میکند. بنابراین، ما از آن دفاع میکنیم که اولاً، ارزیابیهای حرکت ترکیبی از منابع دادههای مکمل را برای ارائه بهترین ارزیابیهای اتصال از نظر عملکردی به کار گیرند. دوم، ما پیشنهاد می کنیم که راهروهای اتصال به دست آمده باید با توجه به نوع حرکت و هندسه مورد بحث قرار گیرند [ 71 ].
3. روند در تعیین کمیت اتصال
ارزیابیهای اتصال یک چارچوب مدلسازی را نشان میدهد که به خوبی برای ارزیابی نقش ساختار چشمانداز در کمک به موفقیت حرکت ارگانیسمها و در تحقق اهداف GI مناسب است [ 72 ]. مطالعات اتصال به طیف وسیعی از گونهها و محیطها از جمله حفاظت از گونههای فردی [ 73 ، 74 ، 75 ]، از جمله بی مهرگان آبزی [ 76 ] تا راهروهای چند گونهای [ 50 ، 57 ، 77 ] تا اجتماعی اعمال شده است. -مطالعات اکولوژیکی در زمینه GI [ 78 ].
گردش کار معمولی یک ارزیابی اتصال زیست محیطی از داده های ورودی زیستی استفاده می کند که از برنامه های نظارتی، آزمایش ها، دانش تخصصی یا از رویکردهای ردیابی منشأ می گیرد. سپس این اطلاعات زیستی به محیط مربوط می شود. محیط با لایههای دادههای فضایی نشان داده میشود که ویژگیهای آنها (عناصر منظر) مقادیری را نشان میدهند که نشاندهنده گرادیان هزینه منظر از مانع تا راهروی مساعد برای ارگانیسمهای مورد مطالعه است که منجر به مقاومت یا سطح هزینه میشود. در میان رویکردها، مسیرهای کمهزینه [ 79 ] یا سطوح کمهزینه [ 38 ، 80] سپس می تواند برای شناسایی مسیرهای حرکتی بالقوه حیوانات در سراسر چشم انداز محاسبه شود. بسیاری از ارزیابیهای ارتباطی فرض میکنند، اما لزوماً بیان نمیکنند که یک حیوان یا دانش کاملی از منظره دارد [ 81 ، 82 ] یا اصلاً حافظه فضایی ندارد (راهپیمایی تصادفی، McRae و همکاران [ 80 ]).
واضح است که شواهدی وجود دارد که عناصر منظر ساختاری یک پروکسی برای اتصال عملکردی هستند [ 45 ، 83 ]. با این حال، همانطور که قبلاً در بخش 2 مشخص شد ، استفاده از چشم انداز به عنوان محرک اصلی محیط ممکن است ساده سازی بیش از حد پیچیدگی اتصال باشد. به عنوان مثال، همانطور که در چندین مثال نشان داده شده است، مناظر ممکن است ذاتاً برای حرکت گونهها و جریان ژن نفوذپذیرتر از حد انتظار باشند [ 12 ، 41 ، 60 ، 62 ، 71 ]. بنابراین، فرآیندهای درون زا (فیزیولوژی، رفتار) حرکت محرک را می توان به طور جامع تری در مطالعات اتصال در نظر گرفت [ 84 ].
روش های زیادی وجود دارد که می تواند برای محاسبه اتصال مورد استفاده قرار گیرد، از محاسبات فاصله زوجی ساده، به عنوان مثال، استفاده از فاصله اقلیدسی تا مسیرهای کم هزینه (LCP) در ArcGIS یا Circuitscape [ 80 ]، یک نرم افزار رایگان برای دانلود. هر دو LCP و Circuitscape مقاومت های متفاوتی از عناصر منظره جداگانه را فرض می کنند که حرکت را تسهیل یا مانع می شود. علیرغم بررسیهای مختلف که بر این واقعیت تاکید میکنند که سطوح مقاومت به مقاومت اختصاص داده شده یا مقادیر هزینه [ 85 ] و پیشنهاداتی در مورد چگونگی بهبود شناسایی مقادیر مقاومت [ 65 ، 86 ]، برخی مطالعات هنوز بر تخصیص خودسرانه مقاومت تکیه دارند. مقادیر، به عنوان مثال [ 87]، در حالی که دیگران ارزش های مقاومت را بر اساس دانش ذینفعان یا متخصصان قرار می دهند [ 68 ، 88 ]. چند مطالعه مقادیر مقاومت را بر اساس (معکوس) زیستگاه مناسب ارزیابی کردند. به عنوان مثال، بنی و همکاران. [ 89 ]، بر ارزیابیهای کمی تکیه کرد، به عنوان مثال، بهینهسازی سطوح مقاومت با استفاده از یک الگوریتم بهینهسازی غیرخطی برای به حداقل رساندن مدل AIC (معیار اطلاعات Akaike) [ 48 ]، تجزیه و تحلیل حساسیت سناریوهای مختلف رقیب ارزش مقاومت [ 90 ]، یا آزمایشهایی برای اختصاص دادن مقادیر مقاومت [ 91]. به دلیل فقدان کلی داده های تجربی در مورد حرکت موجودات، کالیبراسیون سطوح مقاومت و ارزیابی مدل های اتصال یا اعتبار، یک چالش باقی مانده است. با این حال، مطابق با سایر نویسندگان [ 92 ]، ما دو حوزه را می بینیم که می تواند مدل های اتصال را تقویت کرده و آنها را بهتر به بوم شناسی حرکت و اتصال عملکردی پیوند دهد: (1) کالیبراسیون مقادیر مقاومت باید بر سناریوهای مختلف معیارهای مفهومی صحیح تکیه کند و باید در معرض تجزیه و تحلیل حساسیت قرار گیرد [ 65 ، 85 ، 93 ، 94] و (2) خروجی مدل اتصال می تواند مورد ارزیابی یا اعتبارسنجی قرار گیرد (مثلاً خروجی های مدل را با داده های تجربی مقایسه کنید یا رویکردهای مختلف مدل سازی اتصال را مقایسه کنید) تا اطمینان در عملکرد مدل افزایش یابد [ 49 ، 51 ، 95]. علاوه بر این، انجام تحلیلهای حساسیت یا گنجاندن عدم قطعیت در برآورد پارامترها به ویژه برای تحقیقاتی که منجر به توصیههای حفاظتی یا اقدامات حفاظتی میشود، مهم است. احتمالاً، بسیاری از تحقیقاتی که به دنبال برآورد مقاومت هستند، از سطوح مقاومت حاصله در مدلسازی اتصال استفاده میکنند و این اتصالات یا راهروها برای اجرا به برنامهریزان و مدیران زمین ارتقا مییابند. ارائه طیف وسیعی از امکانات برای اقدامات پیشنهادی، شفافیت را به فرآیند میافزاید و احتمال خرید از سوی مدیران زمین و عموم مردم را افزایش میدهد. با این حال، مقالات کمی اثربخشی روش های اتصال را بین یکدیگر مقایسه می کنند (به عنوان مثال LCP و Circuitscape، [ 96 ، 97 )])، یا راهروهای مدل شده را با مشاهدات مقایسه کنید [ 87 ].
همراه با سطوح مقاومتی بهبود یافته از نظر فنی، پیشرفت مفهومی و روش شناختی قابل توجهی، به عنوان مثال، مدل سازی علّی [ 75 ، 98 ]، استحکام و قابلیت اطمینان آنالیزها را به طرق مختلف افزایش داده است، به عنوان مثال، کالیبراسیون سطوح مقاومت [ 65 ، 85 ، 93 ، 94 ] یا مقایسه عملکرد روشهای مختلف اتصال و انواع داده [ 49 ، 51 ، 95]. روشهای مدلسازی پیشرفته یک رویکرد فاکتوریل و چند مدلی را برای ارزیابی فرضیههای جایگزین مانند سطوح مقاومت و شناسایی ترکیبی از عوامل محیطی که از نظر آماری به بهترین شکل توضیح میدهند، بهعنوان مثال، جریان ژن در یک چشمانداز معین را اعمال میکنند [ 50 ، 73 ، 98 ، 99 ، 100 . ]. توسعه دیگر شامل شبیهسازهای اتصال است که بر چالشهای فنی مانند زمانهای محاسباتی طولانی یا بازده الگوریتمی ضعیف غلبه میکنند [ 101 ، 102 ].
4. برنامه ریزی و استراتژی های طراحی برای زیرساخت سبز: رویکردها، ملاحظات، ابزارها
مطالعات اتصال اطلاعات مهمی را برای حفاظت یا برنامهریزی مناطق حفاظتی در زمینههای چشمانداز پویا فضایی فراهم میکند و ابزارهای منطقی برای کمک مؤثر به GI هستند. اولاً، تحقیقات ارتباط ذاتاً بین رشته ای یا حتی فرارشته ای است. رویکردهای بین رشتهای جنبههای به عنوان مثال، بومشناسی جمعیت و حرکت، ژنتیک منظر، جغرافیا و بومشناسی منظر را ترکیب میکنند. در حالی که بومشناسی جمعیت و حرکت، تجزیه و تحلیل را با ویژگیهای خاص گونهای از صفات و رفتار اطلاع میدهد، جغرافیا و بومشناسی منظر، نظریه و دادههای فضایی را برای استخراج مسیرهای حرکتی مناسب در سراسر چشمانداز ارائه میکنند (Leibovici و Claramunt [ 103 ] نظریه بومشناسی منظر را به چارچوب آنتروپی تعمیم میدهند. ، در حالی که پرکل و همکاران [ 104] از تئوری و داده های فضایی برای ارزیابی رشد شهری و تهدیدات اتصال به چشم انداز استفاده کنید). برخی ارزیابیها با گنجاندن کارشناسانی که [ 88 ، 105 ] را کالیبره میکنند یا ورودیها یا خروجیهای مدلسازی شده را ارزیابی میکنند، رویکردهای فرا رشتهای را به تحلیل اتصال اضافه میکنند [ 106 ، 107 ، 108 ]. دوم، خروجیهای مطالعات اتصال اغلب محصولات جذاب بصری هستند که ارتباط بین دانشمندان و پزشکان را بهبود میبخشند [ 109 ]. به عنوان مثال، مدل زیرساخت سبز برای ایالات متحده، ساخته شده توسط Esri، یک شرکت نرم افزار GIS، با همکاری مرکز زیرساخت سبز برای جوامع و تلاش های برنامه ریزی محلی قابل دسترسی است.https://www.esri.com/en-us/industries/green-infrastructure/overview ). نقشههای خروجی مدلهای اتصال امکان انتقال آسان نتایج علمی به مخاطبان غیر علمی را فراهم میکند [ 38 ، 59 ، 110]. بنابراین، ارزیابی های اتصال قوی می تواند نقش مهمی در برنامه های GI داشته باشد. دیدگاه ها و رویکردهای متفاوتی برای در نظر گرفتن GI در برنامه ریزی و طراحی وجود دارد. به نظر می رسد زمینه های مختلف منطقه ای و فرهنگی و همچنین نیازهای انضباطی به مفهوم سازی های مختلف اجرای GI کمک کرده است. به عنوان مثال، از ایالات متحده، دو شاخه متفاوت از تفکر ریشه می گیرند. از یک طرف، نقشهبرداری و تلاش مدلسازی در مقیاس وسیعی وجود دارد که ریشه در بومشناسی منظر و حفاظت دارد، بهکارگیری معیارهای تکه تکهشدن چشمانداز و تنوع زیستی و همچنین مدلهای اتصال برای طراحی، ارزیابی و اولویتبندی برنامههای GI [ 20 ، 111 ، 112 ].]. این مفهوم سازی بر نقشه برداری از راهروهای سبز، هسته های زیستگاه و شاخص های تنوع زیستی تاکید دارد و تا حدی توسط Esri و مرکز زیرساخت سبز [ 113 ] پشتیبانی می شود. https://www.gicinc.org/index.htm ). این نقشهها مجموعه دادههای منطقهای و ملی را گردآوری کردهاند که از یک رویکرد چند سازمانی گردآوری شدهاند، در حالی که ابزارهای GIS مبتنی بر وب در دسترس به برنامهریزان منطقهای و ذینفعان اجازه میدهند تا مناطق را برای GI اولویتبندی کنند ( https://www.esri.com/en-us/industries ) /green-infrastructure/نمای کلی). از سوی دیگر، شهرداریها و برنامهریزان شهری در طرحهای ایجاد تابآوری، بهویژه در زمینه کاهش طوفان و سیل، GI را در نظر میگیرند. در این مورد، GI کمتر از نظر زیستگاه و اتصال، و بیشتر به عنوان یک جایگزین یا مکمل برای زیرساخت خاکستری، با ابزارهای پاسخگویی سطح بسته و ارزیابی برای دولت های محلی [ 114 ، 115 ، 116 ] در نظر گرفته می شود. برخی شروع به مقایسه این ملاحظات متفاوت و امکان سنجی یک برنامه GI برای دستیابی به هر دو هدف کرده اند [ 117 ]. در کاربرد برنامه ریزی GI برای شهر دیترویت، به عنوان مثال، Meerow و Newell [ 118] به این نتیجه رسید که باید بین مکان یابی برای به حداکثر رساندن مدیریت آب طوفان در مقابل اهداف اتصال چشم انداز، معاوضه هایی انجام شود. ژانگ و همکاران [ 111 ] به صراحت سه نوع شناسی طراحی سطح سایت را برای یک مطالعه اعمال کرد.
به همین ترتیب، مدل های اروپایی با چند کارکردی GI دست و پنجه نرم می کنند. زولچوسکا و همکاران [ 117 ]، در واقع، این مفهوم سازی های متفاوت را به عنوان (1) ساختاری، (2) هیدرولوژیکی، و (3) یکپارچه توصیف می کند. استراتژی تنوع زیستی اتحادیه اروپا (GI، Nature 2000) بر شبکه های اکولوژیکی، خدمات اکوسیستم و نیاز به برنامه ریزی فضایی تأکید دارد [ 119 ، 120 ]. با این حال بسیاری برای رویکردهایی برای تطبیق بهتر بینش منطقه ای با اجرای محلی [ 121 ] استدلال می کنند. در یک مطالعه پاناروپایی، Liquete و همکاران. [ 122] رویکردی را برای (1) کمی کردن و نقشهبرداری ظرفیت طبیعی برای خدمات اکوسیستم، و (2) شناسایی زیستگاههای اصلی و دالانهای حیات وحش به کار برد. چندین نویسنده رویکردها یا چارچوب های مفهومی را برای در نظر گرفتن معیارهای چند منظوره در GI پیشنهاد می کنند [ 29 ، 120 ، 123 ]. معمولاً این معیارها شامل معیارهایی مانند: ارزش حفاظتی، ارزش طبیعی، ارزش تفریحی، فرهنگ و/یا برخی از جنبههای رفاه انسان [ 119 ]، و همچنین شناسایی نیازهای شهری [ 123 ] است.
تعدادی از نویسندگان به صراحت به مدل سازی و طراحی اتصال برای GI می پردازند. به عنوان مثال، لیو و همکاران. [ 124 ] از نظریه مدار برای نقشهبرداری از اتصال چشمانداز GI شهری در شهر نانجینگ، چین استفاده کرد. به همین ترتیب، Perkl [ 112 ] پیوندهای چشم انداز را در یک فرآیند طراحی زمین برای راهروهای حیات وحش یکپارچه کرد، و Perkl و همکاران. [ 104 ] تهدید رشد شهری را برای اتصال چشم انداز ارزیابی کرد. در حالی که در دیترویت، ایالات متحده، ژانگ و همکاران. [ 111 ] یک رویکرد LCP را برای نقشهبرداری راهروهای سبز بالقوه که پارکهای شهر را از طریق بستههای خالی و فضاهای سبز کوچک به هم متصل میکند، به عنوان راهی برای برنامهریزی شبکههای GI شهری به کار برد.
5. خیابان ها به سوی ارزیابی های اتصال برای زیرساخت های سبز پایدار آینده
در سال 2012، کنفرانس سازمان ملل متحد در مورد توسعه پایدار Rio+20 با 17 هدف توسعه پایدار (SDGs) ارائه شد که در دستور کار 2030 برای توسعه پایدار [ 125 ] تعریف شد. تنوع زیستی و اکوسیستم و همچنین اقتصاد سبز موضوعات کلیدی SDGs جهانی هستند. این اهداف به چالشها و فرصتهای زیستمحیطی در جهانی تحت سلطه انسانها با اشاره به اکولوژی، فقر، آموزش، برابری جنسیتی و توسعه شهر میپردازد. بحث پایداری، مناطق را تشویق میکند تا سناریوهایی را برای آیندههای مطلوب ایجاد و اجرا کنند که به طور فعال توسط برنامهها و سیاستها شکل میگیرند [ 125 , 126 , 127]. بنابراین، آینده مطلوب برای GI نه تنها بر تداوم تنوع زیستی متمرکز است، بلکه بر چگونگی عملکرد GI می تواند از SDG زیست محیطی، اجتماعی- فرهنگی و اقتصادی حمایت کند ( شکل 3 ).
این مقاله تاکید میکند که ارزیابیهای اتصال کمکهای ارزشمندی برای GI هستند، زیرا هدف آنها درک حرکت گونهها در یک زمینه صریح فضایی است، و رویکرد مبتنی بر داده به کاربران اجازه میدهد تا فرضیههای اتصال را در سراسر مناظر تأیید کنند. با این حال، برای ارتقای استفاده از آنها برای قابلیت اطمینان قوی برنامههای GI، مدلهای اتصال میتوانند از پرداختن به چالشهای زیر سود ببرند: ما پیشنهاد میکنیم در مورد محرکهای مسیرهای اتصال مدلسازیشده با توجه به انواع حرکت و هندسههای مورد استفاده برای شناسایی حرکت و ویژگیهای درونزای دستگاهها فکر کنیم. موجودات تحت مطالعه مطالعاتی که عمدتاً بر ساختار چشمانداز/محیط زیست بهعنوان محرک اصلی حرکت تمرکز میکنند، میتوانند از چارچوبهای حرکتی موجود بهره ببرند که اهمیت نسبی محرکهای برونزا و درونزا را در نظر میگیرند.42 ]. همچنین، ما شفافیت و تفسیر مسیرهای اتصال مدلسازی شده را با توجه به استفاده و محدودیتهای دادههای حرکتی در نظر گرفته تشویق میکنیم. یک رویکرد امیدوارکننده برای کشف استفاده، محدودیت، و هم افزایی بین انواع دادههای مختلف و ارزیابیهای اتصال، به طور ایدهآل بر ترکیبی از منابع داده مکمل تکیه دارد [ 71 ]. همانطور که توسط نویسندگان دیگر [ 65 ، 85 ، 93 ، 94 ] تاکید شده است، ارزیابی ها در مورد استحکام مدل اتصال و حساسیت نسبت به پارامترسازی مدل نیاز به کاوش عمیق بیشتری دارد. استفاده از تکنیک های آماری توصیه شده که مکانیسم های جایگزین را به طور کامل از هم جدا می کند، می تواند به بهبود رویکردهای اتصال نیز کمک کند، به عنوان مثال، [ 128 ]].
ما همچنین چندین راه را می بینیم که در آن اتصال می تواند با استفاده از داده ها و ابزارهای اضافی پیشرفت کند:
- (1)
-
ارزیابی اتصال از داده های منظره سه بعدی مستمر سود می برد. در دهه گذشته شاهد پیشرفت سنجش از دور بودهایم، و ساختار چشمانداز سه بعدی (3D) اکنون در دسترس است [ 129 ، 130 ]، پهپاد (وسایل نقلیه هوایی بدون سرنشین) و حسگرهای فعال از راه دور مانند LiDAR، یک سیستم تشخیص لیزری [ 131 ، 132 ] . شواهد تجربی قانعکننده نشان میدهد که ارزیابیهای اتصال را میتوان با ساختار پوشش گیاهی سهبعدی پیوسته تکمیل کرد تا ویژگیهای چشمانداز مرتبط عملکردی در ارزیابیهای اتصال ارائه شود.
- (2)
-
ارزیابیهای اتصال میتوانند محیطهای پویا را در نظر بگیرند، زیرا رویکردهای ایستا فقط برای تصمیمگیرندگان کاربرد محدودی دارند، با توجه به تغییرات محیطی قابلتوجه مورد انتظار در آینده [ 110 ، 133 ، 134 ، 135 ]. حسابداری برای فرآیندهای پویا مانند شهرنشینی و ارائه پیش بینی ارزیابی های اتصال آینده در یک محیط در حال تغییر ممکن است برای برنامه ریزی GI آینده ارزش زیادی داشته باشد.
- (3)
-
مدلهای مبتنی بر فردی ممکن است تحلیلهای کمهزینه و فاصله هزینه را تکمیل کنند. رویکردهای مبتنی بر فردی ممکن است به مقابله با پیچیدگی حرکت حیوانات کمک کند که توانایی رفتاری و ناوبری آنها محرک های قوی مسیرهای حرکتی تحقق یافته است [ 136 ، 137 ].
- (4)
-
اتصال چند گونه می تواند برای مدیریت حفاظت مهم باشد. در حالی که بیشتر تحلیلهای اتصال بر روی یک گونه متمرکز میشوند، برنامهریزان حفاظت با منابع محدود میتوانند از مدلهایی که الگوهای حرکتی گونههای متعدد را پیشبینی میکنند [ 59 ] سود زیادی ببرند زیرا برای حفاظت و بازسازی کارآمدتر هستند [ 138 ].
- (5)
-
در نهایت، میبینیم که برنامهریزی و طراحی فضایی برای GI هنوز با نحوه ادغام خواستههای چندگانه و ایجاد آشتی بین عملکردهایی که GI در سیستمهای طبیعی و انسانی انجام میدهد، به چالش کشیده میشود. برنامهریزان و طراحان به خوبی میتوانند روی یک دیدگاه و رویکرد یکپارچهتر برای ادغام اهداف چند منظوره تا حد امکان همگرا شوند و از ابزارهایی برای اندازهگیری عملکرد و مبادلات طرحهای فضایی مختلف در تحقق آن اهداف استفاده کنند. در واقع، فرآیندهای طراحی زمین به سرعت در حال پیشرفت هستند تا این نیازها را برآورده کنند [ 139 ]. سپس شهرداری ها و هیئت های حاکمه منطقه ای می توانند از این ابزارهای پشتیبانی تصمیم برای حرکت به سمت اجرای GI استفاده کنند (همانطور که در Stessens و همکاران [ 31 ]).
6. نتیجه گیری
در دهههای اخیر چندین مفهوم برنامهریزی با محوریت حفظ فضاهای سبز و راهروهای مرتبط، از جمله شبکههای اکولوژیکی، زیربخشهای حفاظتی، و زیرساختهای سبز، در میان دیگران [ 117 ] پدیدار شدهاند ( شکل 3 ). تمرکز بیشتر توجه اخیر، زیرساخت سبز شامل داراییهای طبیعی (پارکها، نهرها، دریاچهها، جنگلها و رودخانهها) است که سیستمی از ویژگیهای محیطی به هم پیوسته را برای حفظ فرآیندهای اکولوژیکی و ارائه خدمات اکوسیستمی تشکیل میدهند [ 113 ]. با این حال، چگونه برنامه ریزان و طراحان متوجه خواهند شد که کدام طرح فضایی به بهترین وجه فرآیندهای اکولوژیکی و خدمات اکوسیستمی مورد علاقه را به تصویر می کشد؟
ارزیابی اتصالات یکی از محبوب ترین ابزارهای علمی برای بهینه سازی طراحی راهرو است. به طور کلی، ارزیابیهای اتصال کمکهای ارزشمندی برای درک حرکت گونهها در یک زمینه صریح فضایی هستند زیرا رویکرد مبتنی بر داده اجازه میدهد تا فرضیههای حرکت ارگانیسمی را در سراسر مناظر تأیید کند. با این حال، کاستیهای مفهومی (مثلاً جنبههای بومشناسی حرکت) و کاستیهای روششناختی (مثلاً ارزیابی مقادیر مقاومت و خروجیهای مدل) در حال حاضر استنتاج و تفسیر بومشناختی خروجیهای روش را محدود میکند. ما استدلال می کنیم که مطالعات اتصال باید به صراحت خود را در (1) چارچوبی قرار دهند تا مشخص شود که چه حرکتی با توجه به داده های حرکتی و هندسه کاربردی ارجاع می شود، و در صورت امکان، شامل اطلاعات درون زا برای تکمیل ارزیابی های ساده اتصال مبتنی بر محیط زیست. (2) مقیاس فضا و زمانی که میتواند با دادههای حرکت اعمالشده در نظر گرفته شود تا به طور مؤثر اتصال مربوط به برنامههای حفاظتی را مشخص کند. (3) مطالعات اتصال، که به سطوح مقاومت با اطلاعات تجربی نیاز دارند، از جمله: (4) تجزیه و تحلیل حساسیت و/یا اقدامات ارزیابی برای آزمایش اعتبار نتایج روش. با توجه به پیشرفتهای سریع در سنجش از دور، کاربرد این رویکرد را میتوان با دادههای منظره سه بعدی پیوسته، جنبههای رفتاری و ارزیابیهای چند گونهای که عملکرد ارزیابیهای اتصال را برای کاربرد حفاظتی بهبود میبخشد، گسترش داد. (2) مقیاس فضا و زمانی که میتواند با دادههای حرکت اعمالشده در نظر گرفته شود تا به طور مؤثر اتصال مربوط به برنامههای حفاظتی را مشخص کند. (3) مطالعات اتصال، که به سطوح مقاومت با اطلاعات تجربی نیاز دارند، از جمله: (4) تجزیه و تحلیل حساسیت و/یا اقدامات ارزیابی برای آزمایش اعتبار نتایج روش. با توجه به پیشرفتهای سریع در سنجش از دور، کاربرد این رویکرد را میتوان با دادههای منظره سه بعدی پیوسته، جنبههای رفتاری و ارزیابیهای چند گونهای که عملکرد ارزیابیهای اتصال را برای کاربرد حفاظتی بهبود میبخشد، گسترش داد. (2) مقیاس فضا و زمانی که میتواند با دادههای حرکت اعمالشده در نظر گرفته شود تا به طور مؤثر اتصال مربوط به برنامههای حفاظتی را مشخص کند. (3) مطالعات اتصال، که به سطوح مقاومت با اطلاعات تجربی نیاز دارند، از جمله: (4) تجزیه و تحلیل حساسیت و/یا اقدامات ارزیابی برای آزمایش اعتبار نتایج روش. با توجه به پیشرفتهای سریع در سنجش از دور، کاربرد این رویکرد را میتوان با دادههای منظره سه بعدی پیوسته، جنبههای رفتاری و ارزیابیهای چند گونهای که عملکرد ارزیابیهای اتصال را برای کاربرد حفاظتی بهبود میبخشد، گسترش داد. (4) تجزیه و تحلیل حساسیت و/یا اقدامات ارزیابی برای آزمایش اعتبار نتایج روش. با توجه به پیشرفتهای سریع در سنجش از دور، کاربرد این رویکرد را میتوان با دادههای منظره سه بعدی پیوسته، جنبههای رفتاری و ارزیابیهای چند گونهای که عملکرد ارزیابیهای اتصال را برای کاربرد حفاظتی بهبود میبخشد، گسترش داد. (4) تجزیه و تحلیل حساسیت و/یا اقدامات ارزیابی برای آزمایش اعتبار نتایج روش. با توجه به پیشرفتهای سریع در سنجش از دور، کاربرد این رویکرد را میتوان با دادههای منظره سه بعدی پیوسته، جنبههای رفتاری و ارزیابیهای چند گونهای که عملکرد ارزیابیهای اتصال را برای کاربرد حفاظتی بهبود میبخشد، گسترش داد.
رویکرد زیرساخت سبز دانشمندان، تصمیمگیرندگان و مردم را برای حفظ و پیوند فضاهای باز، حوضههای آب، حیات وحش، زیستگاهها، پارکها و دیگر مناطق طبیعی که کیفیت زندگی، اقتصاد و حس مکان جامعه را غنی و حفظ میکنند، پیشبینی میکند [ 113 ]. ]. با این حال، در حالی که زیرساخت های سبز علاقه زیادی را در بین محققان، متخصصان و تصمیم گیرندگان به خود جلب کرده است، دامنه رویکردها متفاوت بوده و اجرا محدود بوده است. با روششناسی بهبودیافته و منسجم، ارزیابیهای اتصال میتواند به جلو بردن تصمیمات GI کمک کند.
منابع
- اتو، SP سازگاری، گونه زایی و انقراض در آنتروپوسن. Proc. R. Soc. B Biol. علمی 2018 , 285 , 20182047. [ Google Scholar ] [ CrossRef ] [ PubMed ][ نسخه سبز ]
- استفن، دبلیو. برودگیت، دبلیو. دویچ، ال. گافنی، او. لودویگ، سی. مسیر آنتروپوسن: شتاب بزرگ. آنتر Rev. 2015 , 2 , 81-98. [ Google Scholar ] [ CrossRef ]
- ایروین، ای. سمت تاریک نور. Nature 2018 , 553 , 268. [ Google Scholar ] [ CrossRef ] [ PubMed ][ نسخه سبز ]
- کراوس، جی. بومارکو، آر. گواردیولا، ام. Heikkinen، RK; هلم، ا. کوثری، م. لیندبورگ، آر. اوکینگر، ای. پرتل، ام. پینو، جی. و همکاران تکه تکه شدن زیستگاه باعث از دست دادن فوری و با تاخیر زمانی تنوع زیستی در سطوح مختلف تغذیه ای می شود. Ecol. Lett. 2010 ، 12 ، 597-605. [ Google Scholar ]
- Farneda، FZ; گرل، CEV؛ روشا، ر. فریرا، دی اف. لوپز-باسلز، آ. مایر، CFJ پیش بینی از دست دادن تنوع زیستی در اکوسیستم های جزیره و حومه از طریق دریچه جغرافیای زیستی طبقه بندی و عملکردی. اکوگرافی 2019 . [ Google Scholar ] [ CrossRef ]
- فلچر، آر. دیدهم، ر. بانکز-لیته، سی. بارلو، جی. اورز، RM; روزیندل، جی. Holt، RD; گونزالس، آ. پردینی، ر. دامشن، EI; و همکاران آیا تکه تکه شدن زیستگاه برای تنوع زیستی مفید است؟ Biol. حفظ کنید. 2018 ، 226 ، 9-15. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- وربورگ، RW; Osseweijer، F. چارچوبی برای تخمین از دست دادن تنوع زیستی و هزینه های مرتبط به دلیل انتشار نیتروژن از نیروگاه های منفرد. جی. پاک. تولید 2019 , 239 . [ Google Scholar ] [ CrossRef ]
- ساتر، آی. کیناست، اف. بولیگر، جی. زمستان، بی. Pazur, R. تغییرات در تقاضا و عرضه خدمات اکوسیستم تحت سناریوهای استفاده از زمین در آینده در Vorarlberg، اتریش. J. Mt. Sci. 2019 ، در دست چاپ. [ Google Scholar ] [ CrossRef ]
- Fahrig, L. پاسخهای اکولوژیکی به تکه تکه شدن زیستگاه در هر سه. آنو. کشیش اکول. تکامل. سیستم 2017 ، 48 ، 1-23. [ Google Scholar ] [ CrossRef ]
- فهریگ، ال. آریو-رودریگز، وی. بنت، جی. بوچر-لالوند، وی. کازتا، ای. کری، دی جی; Eigenbrod، F. فورد، AT; هریسون، اس پی; جیگر، JAG; و همکاران آیا تکه تکه شدن زیستگاه برای تنوع زیستی مضر است؟ Biol. حفظ کنید. 2019 ، 230 ، 179-186. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- بورکارت، اس. گوگرلی، ف. سن، جی. کوئن، آر. Bolliger، J. ارزیابی عملکرد راهروهای حیات وحش ارزیابی شده توسط متخصص با داده های ژنتیکی: تعیین اولویت ها برای اقدامات مدیریتی در گوزن (Capreolus capreolus) . برنامه پایه Ecol. 2016 ، 17 ، 52-60. [ Google Scholar ] [ CrossRef ]
- لقمان، ح. مولر، آر. ووپل، ا. برادبک، اس. بولیگر، جی. Gugerli، F. هیچ اثر مانع مشخصی از بزرگراه ها و رودخانه گسترده بر ساختار ژنتیکی نیوت آلپ (Ichthyosaura alpestris) در مناظر پر سکونت. حفظ کنید. ژنت 2018 ، 19 ، 673-685. [ Google Scholar ] [ CrossRef ]
- بولیگر، جی. ادواردز، TC; اگنبرگ، اس. اسماعیل، س. سیدل، آی. Kienast، F. متعادل کردن احتمالات بازسازی جنگل و هزینه های نگهداری در مراتع مرتع خشک با اولویت حفاظتی بالا. حفظ کنید. Biol. 2011 ، 25 ، 567-576. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Araujo، MB; کابزا، م. Thuiller، W. هانا، ال. ویلیامز، پی اچ. آیا تغییرات آب و هوایی گونه ها را از ذخایر خارج می کند؟ ارزیابی روشهای انتخاب ذخیره موجود. گلوب. چانگ. Biol. 2004 ، 10 ، 1618-1626. [ Google Scholar ] [ CrossRef ]
- وانگ، جی ایکس؛ بنزاف، ای. به سوی درک بهتر زیرساخت سبز: بررسی انتقادی. Ecol. اندیک. 2018 ، 85 ، 758-772. [ Google Scholar ] [ CrossRef ]
- اسنال، تی. لهتوماکی، جی. آرپونن، آ. الیت، جی. Moilanen، A. طراحی زیرساخت سبز بر اساس اولویتبندی حفاظت فضایی و مدلسازی ویژگیهای تنوع زیستی و خدمات اکوسیستم. محیط زیست مدیریت 2016 ، 57 ، 251-256. [ Google Scholar ]
- کمیسیون اروپایی. زیرساخت سبز (GI)—افزایش سرمایه طبیعی اروپا ؛ EEA: بروکسل، بلژیک، 2013; پ. 149. [ Google Scholar ]
- منطقه اقتصادی اروپا زیرساخت سبز چیست؟ در دسترس آنلاین: https://www.eea.europa.eu/themes/sustainability-transitions/urban-environment/urban-green-infrastructure/what-is-green-infrastructure (در 27 مارس 2020 قابل دسترسی است).
- پریویترا، آر. La Rosa، D. کاهش آسیب پذیری لرزه ای و تقاضای انرژی شهرها از طریق زیرساخت سبز. پایداری 2018 , 10 . [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- لانزاس، ام. هرموسو، وی. دی میگل، اس. بوتا، جی. Brotons, L. طراحی شبکه ای از زیرساخت های سبز برای افزایش ارزش حفاظتی مناطق حفاظت شده و حفظ خدمات اکوسیستم. علمی کل محیط. 2019 ، 651 ، 541-550. [ Google Scholar ] [ CrossRef ]
- برینک، ای. آلدرز، تی. آدام، دی. فلر، آر. هنسلک، ی. هافمن، آ. ایبه، ک. متی دورت، ا. مایر، م. Negrut، NL; و همکاران آبشارهای سبز: مروری بر سازگاری مبتنی بر اکوسیستم در مناطق شهری گلوب. محیط زیست چانگ. هوم ابعاد سیاست. 2016 ، 36 ، 111-123. [ Google Scholar ] [ CrossRef ]
- Derkzen، ML; ون تیفلن، آجا. Verburg، PH زیرساخت سبز برای سازگاری با آب و هوای شهری: دیدگاه ساکنان در مورد تأثیرات آب و هوا و زیرساخت سبز چگونه ترجیحات سازگاری را شکل می دهد؟ Landsc. طرح شهری. 2017 ، 157 ، 106-130. [ Google Scholar ] [ CrossRef ]
- دموزیر، م. اوررو، ک. هایدریش، او. اولازابال، ای. ژنلتی، دی. اوررو، اچ. Bhave، AG; میتال، ن. فلیو، ای. Faehnle، M. کاهش و سازگاری با تغییرات آب و هوایی: ارزیابی چند منظوره و چند مقیاسی زیرساخت شهری سبز. جی. محیط زیست. مدیریت 2014 ، 146 ، 107-115. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- کارلیر، جی. موران، جی. آگنی، تی. Roche, N. اثرات توسعه راه سبز بر اتصال عملکردی خفاش ها. گلوب. Ecol. حفظ کنید. 2019 , 18 , e00613. [ Google Scholar ] [ CrossRef ]
- برتسقی کوچ، سی. اسموند، پ. پیترز، الف. الگوهای فضایی-زمانی در زیرساخت سبز به عنوان محرک تغییر دمای سطح زمین: مورد سیدنی. بین المللی J. Appl. زمین Obs. Geoinf. 2019 , 83 . [ Google Scholar ] [ CrossRef ]
- لیو، XL; وانگ، ی. لی، ی. وو، JS کمی کردن فرآیند مکانی-زمانی شهرنشینی شهرک: یک رویکرد داده محور در مقیاس بزرگ. Isprs Int. J. Geo-Inf. 2019 ، 8 . [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- تیشندورف، ال. Fahrig, L. در مورد استفاده و اندازه گیری اتصال چشم انداز. Oikos 2001 ، 90 ، 7-19. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- Tischendorf، L. آیا شاخص های چشم انداز می توانند فرآیندهای اکولوژیکی را به طور مداوم پیش بینی کنند؟ Landsc. Ecol. 2001 ، 16 ، 235-254. [ Google Scholar ] [ CrossRef ]
- هردالو، آی. تومیک، دی. Pereković، P. اجرای اصول زیرساخت سبز در دوبرونیک، کرواسی برای به حداقل رساندن مشکلات تغییر سیمات. Urbani Izziv 2015 ، 26 ، S38–S49. [ Google Scholar ] [ CrossRef ]
- ماننا، پ. بونفانته، ا. کولاندریا، ام. دی وایو، سی. لانگلا، جی. ماروتا، ال. Mileti، FA; مینیری، ال. وحشتناک، اف. وینگیانی، س. و همکاران یک سیستم پشتیبانی تصمیم گیری جغرافیایی برای کمک به رشد زیتون در مقیاس چشم انداز محاسبه کنید. الکترون. کشاورزی 2020 , 168 . [ Google Scholar ] [ CrossRef ]
- استسنز، پی. خان، AZ; هیزمنز، ام. Canters، F. تجزیه و تحلیل دسترسی و کیفیت فضای سبز شهری: یک مدل مبتنی بر GIS به عنوان پشتیبانی تصمیم گیری فضایی برای خدمات اکوسیستم شهری در بروکسل. اکوسیستم. خدمت 2017 ، 28 ، 328-340. [ Google Scholar ] [ CrossRef ]
- ویلیامز، AE; Worsley-Tonks، KEL؛ Ezenwa، VO محرک ها و پیامدهای تنوع در ارتباط اجتماعی فردی. انیمیشن. رفتار 2017 ، 133 ، 1-9. [ Google Scholar ] [ CrossRef ]
- هولیوک، ام. کاساگراندی، ر. ناتان، آر. ریویلا، ای. اشپیگل، او. روندها و بخش های گمشده در مطالعه اکولوژی حرکت. Proc. Natl. آکادمی علمی ایالات متحده آمریکا 2008 ، 105 ، 19060-19065. [ Google Scholar ] [ CrossRef ] [ PubMed ][ نسخه سبز ]
- لاپوینت، اس. گالری، پ. ویکلسکی، ام. Kays، R. رفتار حیوانات، مدلهای راهرو مبتنی بر هزینه، و راهروهای واقعی. Landsc. Ecol. 2013 ، 28 ، 1615-1630. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- مک کلور، ام ال. هانسن، ای جی; اتصال مدلهای Inman، RM به حرکات: آزمایش پیشبینیهای مدل اتصال در برابر دادههای مهاجرت تجربی و پراکندگی. Landsc. Ecol. 2016 ، 31 ، 1419-1432. [ Google Scholar ] [ CrossRef ]
- جاکیری، جی. بروکت، تی. هیرزل، ا. ساللی، جی. پرین، ن. استنتاج اثرات منظره بر پراکندگی از فواصل ژنتیکی: تا کجا می توانیم پیش برویم؟ مول. Ecol. 2011 ، 29 ، 692-705. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- لی یاو، جی. دیویدسون، ای. McRae، BH; سبز، DM آیا فرآیندهای چشم انداز الگوهای فیلژوگرافی را در قورباغه چوبی پیش بینی می کنند؟ مول. Ecol. 2009 ، 18 ، 1863-1874. [ Google Scholar ] [ CrossRef ]
- دیکسون، بی جی; آلبانو، سی ام. آنانتارامان، ر. بییر، پی. فارگیون، جی. Graves، TA; خاکستری، من؛ هال، KR؛ لاولر، جی جی. لئونارد، PB; و همکاران کاربردهای تئوری مدار در علم اتصال و حفاظت حفظ کنید. Biol. 2019 ، 33 ، 239-249. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- کلارک، RW; براون، WS; استچرت، آر. Zamudio، KR ادغام رفتار فردی و ژنتیک منظره: ساختار جمعیت خواب زمستانی مار زنگی چوبی. مول. Ecol. 2008 ، 17 ، 719-730. [ Google Scholar ] [ CrossRef ]
- Andreasen، AM; استوارت، KM; لانگلند، WS؛ بکمن، جی پی؛ Forister، ML شناسایی دینامیک منبع – سینک در شیرهای کوهستانی حوضه بزرگ. مول. Ecol. 2012 ، 21 ، 5689-5701. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- Reding، DM; کوشمن، SA; Gosselink، TE; کلارک، WR ارتباط رفتار حرکتی و ساختار ژنتیکی در مقیاس خوب برای مدلسازی اتصال چشمانداز برای بابکتها (Lynx rufus) . Landsc. Ecol. 2013 ، 28 ، 471-486. [ Google Scholar ] [ CrossRef ]
- ناتان، آر. Getz، WM; ریویلا، ای. هولیوک، ام. کادمون، ر. سالتز، دی. Smouse، PE یک پارادایم بوم شناسی جنبش برای یکپارچه سازی تحقیقات حرکت ارگانیسمی. Proc. Natl. آکادمی علمی ایالات متحده آمریکا 2008 ، 105 ، 19052-19059. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- کوشمن، SA; رفتار Lewis، JS Movement تمایز ژنتیکی در خرسهای سیاه آمریکایی را توضیح میدهد. Landsc. Ecol. 2010 ، 25 ، 1613-1625. [ Google Scholar ] [ CrossRef ]
- فلچر، آر. یانگ، جی. هوتو، آر. نوسون، ا. روتا، سی. بینش از نظریه اکولوژیکی در دینامیک زمانی و مدلسازی توزیع گونهها. در مدلسازی گونههای پیشبینیکننده و زیستگاه در بومشناسی منظر: مفاهیم و کاربردها . Drew, A., Wiersma, Y., Huettmann, F., Eds. Springer: New York, NY, USA, 2011. [ Google Scholar ] [ CrossRef ]
- فاتبرت، جی. رابینسون، اچ اس. بالمه، جی. اسلوتو، آر. Hunter, L. زیستگاه ساختاری زیستگاه پراکندگی عملکردی یک گوشتخوار بزرگ را پیش بینی می کند: چگونه پلنگ ها لکه ها را تغییر می دهند. Ecol. Appl. 2015 ، 25 ، 1911-1921. [ Google Scholar ] [ CrossRef ] [ PubMed ][ نسخه سبز ]
- باگت، م. ون دایک، اچ. اتصال به چشم انداز و رفتار حیوانات: دانه عملکردی به عنوان یک تعیین کننده کلیدی برای پراکندگی. Landsc. Ecol. 2007 ، 22 ، 1117-1129. [ Google Scholar ] [ CrossRef ]
- نیکسون، ک. سیلبرناگل، جی. پرایس، ج. میلر، ن. Swaty، R. در دسترس بودن زیستگاه برای گونه های پرندگان متعدد تحت سناریوهای حفاظتی جایگزین مدل سازی شده در حوضه آبخیز رودخانه Two Hearted در میشیگان، ایالات متحده آمریکا. جی. نات. حفظ کنید. 2014 ، 22 ، 302-317. [ Google Scholar ] [ CrossRef ]
- پیترمن، ما؛ Connette، جنرال موتورز; سملیچ، RD; سطوح مقاومت اکولوژیکی Eggert، LS، تمایز ژنتیکی در مقیاس ریز را در سمندر جنگلی زمینی پیشبینی میکنند. مول. Ecol. 2014 ، 23 ، 2402-2413. [ Google Scholar ] [ CrossRef ]
- جکسون، CR; مارنویک، ک. لیندزی، PA; روسکفت، ای. رابرتسون، MP ارزیابی روشهای اتصال زیستگاه: مطالعه موردی با سگهای وحشی آفریقایی در معرض خطر در آفریقای جنوبی. Landsc. Ecol. 2016 ، 31 ، 1433-1447. [ Google Scholar ] [ CrossRef ]
- برودی، جی اف. جیوردانو، ای جی؛ دیکسون، بی جی; هبل وایت، م. برنارد، اچ. محدازلان، ج. اندرسون، جی. Ambu، L. ارزیابی اتصال چند گونه چشم انداز در یک جامعه پستانداران استوایی در معرض تهدید. حفظ کنید. Biol. 2015 ، 29 ، 122-132. [ Google Scholar ] [ CrossRef ]
- باند، ML؛ بردلی، سی ام. کیفنر، سی. موریسون، TA; Lee, DE یک رویکرد چند روشی برای ترسیم و اعتبار دهی کریدورهای مهاجرتی. Landsc. Ecol. 2017 ، 32 ، 1705-1721. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- آبراهمز، بی. دی پیترو، دی. گرافیس، ا. Hollander، A. مدیریت تنوع زیستی تحت تغییرات آب و هوا: چالش ها، چارچوب ها و ابزارهای سازگاری. تنوع زیستی حفظ کنید. 2017 ، 26 ، 2277-2293. [ Google Scholar ] [ CrossRef ]
- Lechner, AM; دوئر، وی. هریس، RMB؛ دوئر، ای. Lefroy، EC چارچوبی برای ترکیب رفتار پراکندگی در مقیاس خوب در برنامه ریزی حفاظت از تنوع زیستی. Landsc. طرح شهری. 2015 ، 141 ، 11-23. [ Google Scholar ] [ CrossRef ]
- رید، جی سی. Litvaitis، JA; کالاهان، سی. کارول، RP; Litvaitis، MK; Broman، DJA مدلسازی اتصال چشمانداز برای بابکتها با استفاده از نظر متخصص و مدلهای مشتقشده تجربی: چقدر خوب کار میکنند؟ انیمیشن. حفظ کنید. 2017 ، 20 ، 308-320. [ Google Scholar ] [ CrossRef ]
- چارنی، ND ارزیابی نظر متخصص و مقیاس فضایی در مدل دوزیستان. Ecol. مدل. 2012 ، 242 ، 37-45. [ Google Scholar ] [ CrossRef ]
- میلانسی، پ. هولدرگر، آر. کانیگلیا، آر. فابری، ای. گالاورنی، م. رندی، ای. مدلهای مبتنی بر متخصص در مقابل زیستگاه مناسب برای توسعه سطوح مقاومت در ژنتیک منظر. Oecologia 2017 ، 183 ، 67-79. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- کوئن، EL; بومن، جی. سادووسکی، سی. Walpole، AA اتصال چشم انداز برای حیات وحش: توسعه و اعتبار سنجی نقشه های پیوند چند گونه. روش ها Ecol. تکامل. 2014 ، 5 ، 626-633. [ Google Scholar ] [ CrossRef ]
- شولتز، آ. کلنکه، آر. لوتزه، جی. ووس، م. ویلند، آر. مدلهای زیستگاه ویلکنینگ، بی. در نظریه منظر و مدیریت منابع: پیوند نظریه با عمل . Bissonette, J., Storch, I., Eds. مطبوعات جزیره: واشنگتن، دی سی، ایالات متحده آمریکا، 2003; ص 261-282. [ Google Scholar ]
- کلر، دی. هولدرگر، آر. ون استرین، ام جی; Bolliger, J. چگونه می توان ژنتیک چشم انداز را برای مدیریت حفاظت مفید ساخت؟ حفظ کنید. ژنت 2015 ، 16 ، 503-512. [ Google Scholar ] [ CrossRef ]
- ون استرین، ام جی; کلر، دی. هولدرگر، آر. غزول، ج. کیناست، اف. بولیگر، جی. ژنتیک منظر به عنوان ابزاری برای برنامه ریزی حفاظت: پیش بینی اثرات تغییر چشم انداز بر جریان ژن. Ecol. Appl. 2014 ، 24 ، 327-339. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- بولیگر، جی. کلر، دی. هولدرگر، R. وقتی متغیرهای چشمانداز نرخ مهاجرت را توضیح نمیدهند: نمونهای از سنجاقک در معرض خطر (Leucorrhinia caudalis) . یورو J. Entomol. 2011 ، 108 ، 327-330. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- لی لی، جی. آنجلون، اس. فلوری، سی. هولدرگر، آر. Bolliger, J. افزایش تراکم برکه برای حفظ یک شبکه زیستگاه تکه تکه قورباغه درختی اروپایی (Hyla arborea) . جی. هرپتول. 2015 ، 49 ، 217-221. [ Google Scholar ] [ CrossRef ]
- فایفر، وی. سیلبرناگل، جی. گودوت، سی. Zalapa، J. Woodland و غنای گل، تراکم زنبورهای بامبل را در مناظر پالس منابع زغال اخته افزایش می دهد. Landsc. Ecol. 2019 ، 34 ، 979–996. [ Google Scholar ] [ CrossRef ]
- ردیف، JR؛ بلوین-دمرز، جی. توزیع زیستگاه Lougheed، SC بر پراکندگی و ساختار جمعیت ژنتیکی در مقیاس ریز مارهای روباه شرقی (Mintonius gloydi) در سراسر یک چشمانداز تکه تکه تأثیر میگذارد. مول. Ecol. 2010 ، 19 ، 5157-5171. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- زلر، کالیفرنیا؛ مک گریگال، ک. Whiteley، AR برآورد مقاومت منظره در برابر حرکت: یک بررسی. Landsc. Ecol. 2012 ، 27 ، 777-797. [ Google Scholar ] [ CrossRef ]
- یومنام، بی. Jhala، YV; قریشی، ق. مالدونادو، جی. گوپال، آر. ساینی، س. سرینیواس، ی. Fleischer, RC اولویت دادن به حفاظت از ببر از طریق ژنتیک چشم انداز و پیوندهای زیستگاهی. PLoS ONE 2014 ، 9 ، e111207. [ Google Scholar ] [ CrossRef ] [ PubMed ][ نسخه سبز ]
- پریچارد، جی کی; استفنز، ام. دانلی، ص. استنتاج ساختار جمعیت با استفاده از داده های ژنوتیپ چند لوکوس. ژنتیک 2000 ، 155 ، 945-959. [ Google Scholar ] [ PubMed ]
- گریزلز، اس. د بلوک، جی جی; مکوندی، ر. وانمچلن، ک. بروکهو، جی. مازوچ، وی. سامبرا، آر. زیما، ج. لیرز، اچ. Baird، SJE تمایز ژنتیکی بین موشهای چند پستاندار شهری و روستایی به هم پیوسته در تانزانیا علیرغم جریان ژن. J. Evol. Biol. 2016 ، 29 ، 1952-1967. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- هریسون، کالیفرنیا؛ پاولوا، آ. آموس، JN; رادفورد، جی کیو؛ Sunnucks، P. آیا کاهش تحرک از طریق مناظر تکه تکه شده، الگوهای انقراض تکهای را برای سه عسلخوار توضیح میدهد؟ J. Anim. Ecol. 2014 ، 83 ، 616-627. [ Google Scholar ] [ CrossRef ]
- هریسون، کالیفرنیا؛ پاولوا، آ. آموس، JN; تاکوچی، ن. لیل، آ. رادفورد، جی کیو؛ Sunnucks، P. فرآیندهای جمعیتی در مقیاس ریز را در مناظر تکهتکهشده با وجود اتصال ژنتیکی در مقیاس بزرگ برای یک پرورشدهنده تعاونی گسترده و مشترک مختل کرد: پری-ورن عالی (Malurus cyaneus) . J. Anim. Ecol. 2013 ، 82 ، 322-333. [ Google Scholar ] [ CrossRef ]
- فری، م. Csencsics، C.; برادبک، اس. شوایزر، ای. بوهلر، سی. گوگرلی، ف. Bolliger، J. ترکیب ژنتیک منظر، ردیابی رادیویی و نظارت طولانی مدت برای استخراج پیامدهای مدیریتی برای وزغ Natterjack (Epidalea calamita) در مناظر کشاورزی. جی. نات. حفظ کنید. 2016 ، 32 ، 22-34. [ Google Scholar ] [ CrossRef ]
- نایدو، ر. کیلیان، جی دبلیو. دو پریز، پی. بیتل، پی. آسشنبورن، او. تیلور، RD; استوارت هیل، جی. ارزیابی اثربخشی راهروهای حیات وحش در مقیاس محلی و منطقه ای با استفاده از معیارهای کمی اتصال عملکردی. Biol. حفظ کنید. 2018 ، 217 ، 96-103. [ Google Scholar ] [ CrossRef ]
- Wasserman، TN; کوشمن، SA; شوارتز، MK; Wallin، DO مقیاس بندی فضایی و استنتاج چند مدل در ژنتیک منظر: Martes americana در شمال آیداهو. Landsc. Ecol. 2010 ، 25 ، 1601-1612. [ Google Scholar ] [ CrossRef ]
- اسکوایرز، جی آر. DeCesare، NJ; اولسون، LE; Kolbe، JA; هبل وایت، م. Parks, SA ترکیبی از انتخاب منبع و رفتار حرکتی برای پیشبینی راهروها برای سیاهگوش کانادا در حاشیه محدوده جنوبی آنها. Biol. حفظ کنید. 2013 ، 157 ، 187-195. [ Google Scholar ] [ CrossRef ]
- پارک ها، ال سی. والین، DO; کوشمن، SA; McRae، BH تجزیه و تحلیل سطح چشم انداز اتصال جمعیت بزهای کوهی در واشنگتن و جنوب بریتیش کلمبیا. حفظ کنید. ژنت 2015 ، 16 ، 1195-1207. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- موران-اوردونز، آ. پاولوا، آ. پیندر، AM; سیم، ال. Sunnucks، P. تامپسون، آر.ام. دیویس، جی. جوامع آبزی در مناظر خشک: شرایط محلی، صفات پراکندگی و پیکربندی چشم انداز تنوع زیستی محلی را تعیین می کند. غواصان. توزیع کنید. 2015 ، 21 ، 1230-1241. [ Google Scholar ] [ CrossRef ]
- بلیله، بی. باومن، ام. گریفیث، پی. هایدلبرگ، ا. مانولیان، ک. Radeloff، VC; زازناشویلی، ن. Kuemmerle، T. ارزیابی اتصال چشم انداز برای پستانداران بزرگ در قفقاز با استفاده از ترکیبات تصویر فصلی Landsat 8. سنسور از راه دور محیط. 2017 ، 193 ، 193-203. [ Google Scholar ] [ CrossRef ]
- شیو، ن. ایگناتیوا، م. ون دن بوش، CK; چای، YY; وانگ، اف. Cui، TF; یانگ، FP دیدگاه اجتماعی-اکولوژیکی شبکه های سبز شهری: مورد استکهلم. اکوسیست شهری. 2017 ، 20 ، 729-742. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- آدریانسن، اف. Chardon، JP; دی بلوست، جی. سوئینن، ای. ویلالبا، اس. گلنیک، اچ. ماتیسن، ای. کاربرد مدلسازی «کمترین هزینه» به عنوان مدلهای منظر عملکردی. Landsc. طرح شهری. 2003 ، 64 ، 233-247. [ Google Scholar ] [ CrossRef ]
- McRae، BH; دیکسون، بی جی; کیت، تی. شاه، VB با استفاده از نظریه مدار برای مدل سازی اتصال در محیط زیست و حفاظت. اکولوژی 2008 ، 10 ، 2712-2724. [ Google Scholar ] [ CrossRef ]
- مویلانن، ا. ویلسون، کالیفرنیا؛ پوسینگام، اچ. اولویتبندی حفاظت فضایی: روشهای کمی و ابزارهای محاسباتی . انتشارات دانشگاه: آکسفورد، انگلستان، 2009. [ Google Scholar ]
- پینتو، ن. کیت، TH فراتر از مسیر کمهزینه: ارزیابی افزونگی راهرو با استفاده از رویکرد نظری گراف. Landsc. Ecol. 2009 ، 24 ، 253-266. [ Google Scholar ] [ CrossRef ]
- فاتبرت، جی. بابت، ای. اسلوتو، آر. فیشر، سی. اثرات چشم انداز بر اندازه محدوده خانه گراز وحشی تحت رژیم های برداشت متضاد در یک اکوسیستم کشاورزی تحت سلطه انسان. یورو جی. وایلدل. Res. 2017 , 63 . [ Google Scholar ] [ CrossRef ]
- بولیگر، جی. لندر، تی. بالکنهول، N. ژنتیک چشم انداز از سال 2003: وضعیت، چالش ها و جهت گیری های آینده. Landsc. Ecol. 2014 ، 29 ، 361-366. [ Google Scholar ] [ CrossRef ]
- ریفیلد، بی. فورتین، ام جی. Fall، A. حساسیت نمودارهای زیستگاه کم هزینه به مقادیر سطح هزینه نسبی. Landsc. Ecol. 2010 ، 25 ، 519-532. [ Google Scholar ] [ CrossRef ]
- پانزاکی، ام. ون مورتر، بی. استرند، او. Saerens، M. کی، آی کی؛ سنت کلر، سی سی; هرفیندال، آی. Boitani, L. پیشبینی پیوستگی بین راهروها و موانع برای حرکت حیوانات با استفاده از توابع انتخاب مرحله و کوتاهترین مسیرهای تصادفی. J. Anim. Ecol. 2016 ، 85 ، 32-42. [ Google Scholar ] [ CrossRef ] [ PubMed ][ نسخه سبز ]
- Grafius، DR; کورستانژ، ر. Siriwardena، GM; پلامر، KE; هریس، JA یک نمای چشم پرنده: استفاده از نظریه مدار برای مطالعه اتصال منظر شهری برای پرندگان. Landsc. Ecol. 2017 ، 32 ، 1771-1787. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- Lechner, AM; اسپرود، دی. کارتر، او. Lefroy، EC مشخص کردن اتصال چشمانداز برای برنامهریزی حفاظت با استفاده از رویکرد صنفی پراکندگی. Landsc. Ecol. 2017 ، 32 ، 99-113. [ Google Scholar ] [ CrossRef ]
- بنی، ل. پیزا، جی. لوپی، م. اسپیلوتروس، جی. فابری، ای. رندی، ای. Orioli، V. ارزیابی اتصال اکولوژیکی در جمعیت سمندر آتشی با ساختار قوی (Salamandra salamandra) . Ecol. تکامل. 2015 ، 5 ، 3472-3485. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- بریکر، اس. مورتی، ام. بوش، آر. غزول، ج. Obrist، MK; Bontadina، F. ارزیابی اتصال زیستگاه برای حیوانات ساکن زمین در یک محیط شهری. Ecol. Appl. 2014 ، 24 ، 1583-1595. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- نواکوفسکی، ای جی. ویمن-اچوریا، ام. کرز، دی جی; Donnelly، MA ارزیابی اتصال برای دوزیستان استوایی با استفاده از سطوح مقاومت تجربی مشتق شده. Ecol. Appl. 2015 ، 25 ، 928-942. [ Google Scholar ] [ CrossRef ]
- زلر، کالیفرنیا؛ جنینگز، MK; ویکرز، TW; ارنست، HB; کوشمن، SA; Boyce، WM آیا همه انواع داده ها و مدل های اتصال برابر هستند؟ اعتبارسنجی رویکردهای اتصال رایج با داده های پراکندگی غواصان. توزیع کنید. 2018 ، 24 ، 868-879. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- گریوز، تی. چندلر، RB; رویل، جی. بییر، پی. کندال، KC برآورد مقاومت چشم انداز در برابر پراکندگی. Landsc. Ecol. 2014 ، 29 ، 1201-1211. [ Google Scholar ] [ CrossRef ]
- کوئن، EL; بومن، جی. Walpole, AA اثر پارامترسازی سطح هزینه بر برآورد مقاومت منظر. مول. Ecol. منبع. 2012 ، 12 ، 686-696. [ Google Scholar ] [ CrossRef ]
- Dilts، TE; ویزبرگ، پی جی. لایتنر، پی. Matocq، MD; Inman، RD; Nussear، KE; Esque، اتصال چند مقیاسی TC و نظریه گراف، مناطق حیاتی را برای حفاظت تحت تغییرات آب و هوا برجسته می کند. Ecol. Appl. 2016 ، 26 ، 1223-1237. [ Google Scholar ] [ CrossRef ]
- گاریدو-گاردونو، تی. تلز-والدز، او. مانل، اس. Vazquez-Dominguez، E. نقش ناهمگونی زیستگاه و اتصال چشم انداز در شکل دادن به جریان ژن و ساختار جمعیت فضایی یک گونه جونده غالب در یک جنگل خشک استوایی. جی. زول. 2016 ، 298 ، 293-302. [ Google Scholar ] [ CrossRef ]
- چورکو، جی. کیناست، اف. Bolliger، J. ارزیابی چند گونه برای شناسایی اتصال عملکردی در یک چشم انداز تحت سلطه انسان. بین المللی جی. جئوگر. Inf. سیستم 2020 ، در دست چاپ. [ Google Scholar ]
- کوشمن، SA; McKelvey، KS; هیدن، جی. شوارتز، جریان ژن MK در مناظر پیچیده: آزمایش فرضیه های متعدد با مدل سازی علی صبح. نات. 2006 ، 168 ، 486-499. [ Google Scholar ] [ CrossRef ] [ PubMed ][ نسخه سبز ]
- ردی، PA; کوشمن، SA; سریواستاوا، ا. سرکار، ام اس; Shivaji، S. Tiger فراوانی و جریان ژن در هند مرکزی توسط ترکیبات متفاوت توپوگرافی و پوشش زمین هدایت می شود. غواصان. توزیع کنید. 2017 ، 23 ، 863-874. [ Google Scholar ] [ CrossRef ]
- ورگارا، م. کوشمن، SA; رویز-گونزالس، A. تفاوت های اکولوژیکی و عوامل محدود کننده در زمینه های مختلف منطقه ای: ژنتیک چشم انداز ماتن سنگی در شبه جزیره ایبری. Landsc. Ecol. 2017 ، 32 ، 1269-1283. [ Google Scholar ] [ CrossRef ]
- Landguth، EL; دست، BK; گلسی، جی. کوشمن، SA; Sawaya, MA UNICOR: یک شبیهساز شبکه ارتباطی و راهروی گونهها. اکوگرافی 2012 ، 35 ، 9-14. [ Google Scholar ] [ CrossRef ]
- کوئن، EL; الینگتون، ای اچ. Bowman, J. نقشهبرداری اتصال چشمانداز برای گسترههای فضایی بزرگ. Landsc. Ecol. 2019 ، 34 ، 2421-2433. [ Google Scholar ] [ CrossRef ]
- Leibovici، DG; Claraunt, C. در مورد ادغام توزیعهای اندازه و شکل در چارچوب آنتروپی اطلاعات مکانی-زمانی. آنتروپی 2019 ، 21 . [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- پرکل، آر. نورمن، ال.ام. میچل، دی. فلر، ام. اسمیت، جی. ارزیابی تهدیدات رشد شهری و اتصال منظر ویلسون، NR در پارک ملی ساگوارو، آریزونا، ایالات متحده. J. کاربری زمین علمی. 2018 ، 13 ، 102-117. [ Google Scholar ] [ CrossRef ]
- کراسبی، ام. برکهایمر، آی. پیرس، دی جی; Singleton، PH; هال، SA; هالوپکا، KC; Gaines، WL; لانگ، RA; McRae، BH; کوسنتینو، BL; و همکاران مدلهای راهروی گونههای کانونی و «طبیعی بودن» منظر، رویکردهای مکملی را برای برنامهریزی حفاظت از اتصال ارائه میدهند. Landsc. Ecol. 2015 ، 30 ، 2121-2132. [ Google Scholar ] [ CrossRef ]
- فریمن، سی اف; Bell، KP Conservation در مقابل تقسیمبندیهای خوشهای و مفاهیم برای اتصال زیستگاه. Landsc. طرح شهری. 2011 ، 10 ، 30-42. [ Google Scholar ] [ CrossRef ]
- Sawyer, SC; Epps، CW; Brashares، JS قرار دادن پیوندها در میان زیستگاه های تکه تکه شده: آیا مدل های کم هزینه نشان دهنده نحوه استفاده حیوانات از مناظر هستند؟ J. Appl. Ecol. 2011 ، 48 ، 668-678. [ Google Scholar ] [ CrossRef ]
- Marrotte, RR; بومن، جی. براون، MGC؛ کوردس، سی. موریس، کی. پرنتیس، مگابایت؛ Wilson، PJ اتصال ژنتیکی چند گونه در یک شبکه زیستگاه زمینی. حرکت Ecol. 2017 ، 5 . [ Google Scholar ] [ CrossRef ] [ PubMed ][ نسخه سبز ]
- ووکومانوویچ، جی. اسکریپ، م. Meentenmeyer، R. ساخت آن به صورت فضایی، آن را شخصی می کند: درگیر کردن سهامداران با مدل سازی مشارکتی جغرافیایی. سرزمین 2019 ، 8 ، 38. [ Google Scholar ] [ CrossRef ] [ نسخه سبز ]
- Lechner, AM; هریس، RMS؛ دوئر، وی. دوئر، ای. دریلسما، ام. Lefroy، EC از مدلسازی اتصال استاتیک تا برنامهریزی مبتنی بر سناریو در مقیاسهای محلی و منطقهای. جی. نات. حفظ کنید. 2015 ، 28 ، 78-88. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- ژانگ، ز. میرو، اس. نیول، جی. لیندکوئیست، ام. افزایش اتصال چشم انداز از طریق مدل سازی و طراحی راهرو زیرساخت سبز چند منظوره. شهری برای. سبز شهری. 2019 ، 38 ، 305-317. [ Google Scholar ] [ CrossRef ]
- Perkl, RM Geodesigning Links Landscape: Couping GIS با طراحی کریدور حیات وحش در برنامه ریزی حفاظت. Landsc. طرح شهری. 2016 ، 156 ، 44-58. [ Google Scholar ] [ CrossRef ]
- Firehock، KE; Walker, RA Green Infrastructure: نقشه و برنامه ریزی جهان طبیعی با GIS . Esri Press: Redlands، CA، USA، 2019. [ Google Scholar ]
- ویسکانسین، جی. مقابله با موانع زیرساخت سبز: ممیزی قوانین و مقررات محلی . WI، ایالات متحده آمریکا، 2017; در دسترس آنلاین: https://www.seagrant.wisc.edu/our-work/focus-areas/coastal-communities/green-infrastructure/ (در 1 ژانویه 2020 قابل دسترسی است).
- بندیکت، ام. مک ماهون، ای. برگن، ال. زیرساخت سبز: پیوند مناظر و جوامع . مطبوعات جزیره: واشنگتن، دی سی، ایالات متحده آمریکا، 2006. [ Google Scholar ]
- لینچ، الف. آیا سبز بودن خوب است؟ ارزیابی نتایج اکولوژیکی برنامه ریزی زیرساخت سبز شهرستان. جی. پلان. آموزش. Res. 2016 ، 36 ، 90-104. [ Google Scholar ] [ CrossRef ]
- سولچوسکا، بی. گیدیچ، آر. آیا میتوانیم با این چالش روبرو شویم: چگونه مفهوم نظری زیرساخت سبز را در عمل برنامهریزی پیادهسازی کنیم؟ مطالعه موردی ورشو Landsc. Res. 2017 ، 42 ، 76-194. [ Google Scholar ] [ CrossRef ]
- میرو، اس. نیوول، جی. برنامه ریزی فضایی برای زیرساخت سبز چند منظوره: انعطاف پذیری در حال رشد در دیترویت. Landsc. طرح شهری. 2017 ، 159 ، 62-75. [ Google Scholar ] [ CrossRef ]
- لای، اس. لئون، F. پل زدن اهداف حفاظت از تنوع زیستی با برنامه ریزی منظر از طریق زیرساخت های سبز: مطالعه موردی از ساردینیا، ایتالیا. در یادداشت های سخنرانی در علوم کامپیوتر، مجموعه مقالات علوم محاسباتی و کاربردهای آن، تریست، ایتالیا، 3 تا 6 ژوئیه 2017 . Springer: Cham, Switzerland, 2017; جلد 10409، ص 10456–10472. [ Google Scholar ]
- لافورتزا، آر. دیویس، سی. سانسی، جی. Konijnendijk، C. زیرساخت سبز به عنوان ابزاری برای حمایت از برنامه ریزی فضایی در مناطق شهری اروپا. iForest Biogeosci. برای. 2013 ، 6 ، 102-108. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- ریمر، ام. Rusche، K. زیرساخت سبز تحت فشار. روایتی جهانی بین چشم انداز منطقه ای و اجرای محلی یورو طرح. گل میخ. 2019 ، 27 ، 1542-1563. [ Google Scholar ] [ CrossRef ]
- لیکت، سی. کلیشولت، اس. دیج، جی. میس، جی. گریزتی، بی. اولاه، بی. زولیان، جی. نقشه برداری زیرساخت سبز بر اساس خدمات اکوسیستم و شبکه های اکولوژیکی: مطالعه موردی پان-اروپایی. محیط زیست علمی سیاست 2015 ، 54 ، 268-280. [ Google Scholar ] [ CrossRef ]
- واسیلیویچ، ن. رادیک، بی. گاوریلوویچ، اس. اسلوکیچ، بی. مدارویچ، م. مفهوم زیرساخت سبز و برنامه ریزی منظر شهری: چالشی برای برنامه ریزی جنگلداری شهری در بلگراد، صربستان. iForest Biogeosci. برای. 2018 ، 11 ، 491-498. [ Google Scholar ] [ CrossRef ]
- لیو، اس. یین، ی. لی، جی. چنگ، اف. دونگ، اس. Zhang، Y. استفاده از شاخصهای اتصال چشمانداز در مقیاس متقابل برای شناسایی تکههای منابع زیستگاهی کلیدی برای فیلهای آسیایی در Xishuangbanna، چین. Landsc. طرح شهری. 2018 ، 171 ، 80-87. [ Google Scholar ] [ CrossRef ]
- سازمان ملل. درباره اهداف توسعه پایدار در دسترس آنلاین: https://www.un.org/sustainabledevelopment/sustainable-development-goals/ (دسترسی در 15 مه 2009).
- بای، XM; ون در لیو، اس. اوبراین، ک. برخوت، ف. بیرمن، اف. Brondizio، ES; کودنک، سی. عزیز، جی. دورایاپا، ع. گلیزر، ام. و همکاران آینده های قابل قبول و مطلوب در آنتروپوسن: دستور کار تحقیقاتی جدید گلوب. محیط زیست چانگ. هوم ابعاد سیاست. 2016 ، 39 ، 351-362. [ Google Scholar ] [ CrossRef ]
- زمین آینده دستور کار تحقیق. 2014. در دسترس آنلاین: https://www.futureearth.org/sites/default/files/strategic_research_agenda_2014.pdf (در 27 مارس 2020 قابل دسترسی است).
- ون استرین، ام جی; کلر، دی. هولدرگر، آر. یک رویکرد تحلیلی جدید برای مدلسازی ژنتیکی منظر: تحلیل ترانسکت کمهزینه و مدلهای مختلط خطی. مول. Ecol. 2012 ، 21 ، 4010-4023. [ Google Scholar ] [ CrossRef ]
- گینزلر، سی. Hobi, ML تطبیق تصویر استریو در سراسر کشور برای بهروزرسانی مدلهای سطح دیجیتال در چارچوب فهرست فهرست جنگلهای ملی سوئیس. Remote Sens. 2015 ، 7 ، 4343-4370. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- راندین، CF; اشکرافت، ام. بولیگر، جی. کاوندر-بارز، جی. کوپس، ن. دالینگر، اس. دیرنبوک، تی. اکرت، اس. الیس، ای. جولیانی، جی. و همکاران به سوی بهبود نظارت بر اکوسیستمهای زمینی در Anthropocene از طریق ادغام نزدیکتر دادههای سنجش از راه دور در مدلهای توزیع گونهها. سنسور از راه دور محیط. 2020 ، در دست چاپ. [ Google Scholar ]
- برگن، KM; گوتز، اس جی. دبیه، RO; Henebry، جنرال موتورز; هونساکر، سی تی. ایمهوف، ام ال. نلسون، RF؛ پارکر، جی جی؛ Radeloff، VC سنجش از دور ساختار سه بعدی پوشش گیاهی برای تنوع زیستی و زیستگاه: بررسی و مفاهیم برای ماموریت های فضایی لیدار و رادار. جی. ژئوفیز. Res. 2009 ، 114 ، G00E06. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- مریک، ام جی; Koprowski، نظریه مدار JL برای تخمین مسیرهای پراکندگی زایمان و اتصال عملکردی چشم انداز برای یک پستاندار کوچک در معرض خطر. Landsc. Ecol. 2017 ، 32 ، 1163-1179. [ Google Scholar ] [ CrossRef ]
- مگینی، ر. لمان، ا. زبیندن، ن. Zimmermann، NE; بولیگر، جی. شرودر، بی. فوپن، آر. اشمید، اچ. بنیستون، ام. جنی، ال. ارزیابی آسیب پذیری گونه ها در برابر تغییرات آب و هوا و کاربری زمین: مورد پرندگان پرورش دهنده سوئیس. غواصان. توزیع کنید. 2014 ، 20 ، 708-719. [ Google Scholar ] [ CrossRef ]
- لئونارد، پی. ساترلند، آر. بالدوین، آر. فدک، د. کارنس، آر. مونتگومری، الف. تلفات اتصال چشم انداز به دلیل افزایش سطح دریا و تغییر کاربری زمین. انیمیشن. حفظ کنید. 2017 ، 20 ، 80-90. [ Google Scholar ] [ CrossRef ]
- نه، ANM; کورستانژ، ر. هریس، جی. Grafius، DR; Siriwardena، شبکه های ارتباطی زیست محیطی GM در شهرهایی که به سرعت در حال گسترش هستند. Heliyon 2017 , 3 . [ Google Scholar ] [ CrossRef ] [ PubMed ][ نسخه سبز ]
- كناگارج، ر. ویگاند، تی. کرامر-شادت، اس. Goyal، SP استفاده از مدلهای حرکتی مبتنی بر فردی برای ارزیابی اتصال بین وصلهای برای گوشتخواران بزرگ در مناظر تکه تکه شده. Biol. حفظ کنید. 2013 ، 167 ، 298-309. [ Google Scholar ] [ CrossRef ]
- پولانسکی، ال. کیلیان، دبلیو. Wittemyer، G. توضیح اهمیت حافظه فضایی در تصمیم گیری های حرکتی توسط فیل های ساوانای آفریقایی با استفاده از مدل های حالت-فضا. Proc. R. Soc. B Biol. علمی 2015 ، 282 ، 20143042. [ Google Scholar ] [ CrossRef ] [ نسخه سبز ]
- فلیشمن، ای. اندرسون، جی. مدلهای اتصال تکگونهای و چند گونهای Dickson، BG برای پستانداران بزرگ در کشور ناواهو. غرب. N. Am. نات. 2017 ، 77 ، 237-251. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- رودریگز-اسپینوس، VM؛ آگیلرا-بناونته، اف. Gimez-Delgado، M. طراحی زیرساخت سبز با استفاده از GIS و تجزیه و تحلیل فضایی: پیشنهادی برای کریدور هنارس (مادرید-گوادالاخارا، اسپانیا). Landsc. Res. 2020 ، 45 ، 26-43. [ Google Scholar ] [ CrossRef ]

شکل 1. تغییرات جهانی محیطی و فشار انسانی نیازمند برنامه ریزی و طراحی زیرساخت سبز (GI) برای اطمینان از تداوم طولانی مدت اتصال عملکردی در مقیاس چشم انداز در مناطق تحت تسلط انسان است.
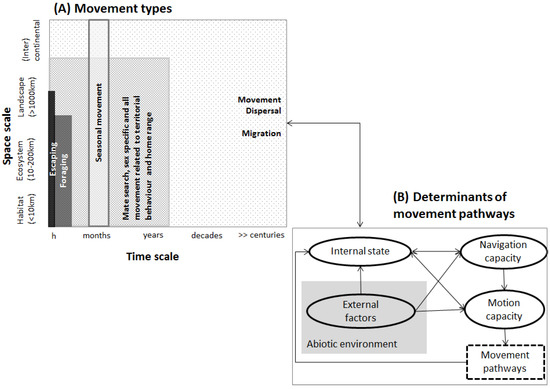
شکل 2. ( الف ) انواع حرکت و ( B ) عوامل تعیین کننده مسیرهای حرکت (ساده شده از ناتان و همکاران [ 42 ]).
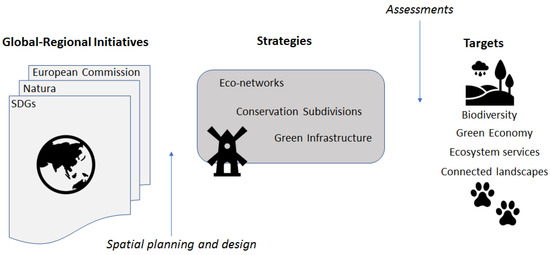
شکل 3. رابطه مفهومی بین ابتکارات بین المللی در مقیاس وسیع برای دستیابی به اهداف در اهداف انسان-محیط (به عنوان مثال مناظر مرتبط) از طریق استراتژی های مختلف مبتنی بر منظر. برنامه ریزی و طراحی فضایی برنامه هایی را برای این استراتژی ها از جمله زیرساخت سبز تولید می کند، در حالی که ارزیابی های مختلف از جمله مدل های اتصال، استراتژی ها را در دستیابی به اهداف اطلاع رسانی و ارزیابی می کند. اهداف توسعه پایدار = اهداف توسعه پایدار جهانی.


بدون دیدگاه