

چکیده
علف مارماهی کوتوله Zostera noltei Hornemann ( Z. noltei ) غالب ترین علف دریایی در سیستم های ساحلی نیمه محصور در سواحل اقیانوس اطلس مراکش است. این گونه در سرتاسر جهان در حال کاهش است و نظارت بر وسعت مراتع آن یک رویکرد مفید برای برآورد اثرات عوامل استرس زای طبیعی و انسانی خواهد بود. در اینجا، هدف ما ترسیم نقشه Z. noltei بودمراتع در تالاب ساحلی Merja Zerga (ساحل اقیانوس اطلس مراکش) با استفاده از سنجش از دور. ما از یک الگوریتم جنگل تصادفی ترکیب شده با داده های میدانی برای طبقه بندی یک تصویر ماهواره ای SPOT 7 استفاده کردیم. علیرغم مشکلات مربوط به عدم همگام سازی تصاویر ماهواره ای با ضریب جزر و مد بالا، نتایج ما با دقت 95 درصد نشان داد که بسترهای علف مارماهی کوتوله را می توان با موفقیت از سایر زیستگاه ها در تالاب متمایز کرد. مساحت تخمین زده شده با در نظر گرفتن بسترهای مختلط ( Z. noltei-associated macroalgae ) 160.76 هکتار بود. به نظر می رسد استفاده از تصاویر ماهواره ای SPOT 7 برای نظارت طولانی مدت Z. noltei رضایت بخش باشد.مراتع در تالاب Merja Zerga و برای تخمین زیست توده با استفاده از یک رابطه کمی NDVI-زیست توده. با این وجود، استفاده از این روش تخمین زیست توده برای مراتع علف مارماهی کوتوله در مناطقی که NDVI به دلیل انباشته شدن چندین لایه اشباع شده است، می تواند ناموفق باشد.
کلید واژه ها:
علف دریایی ; سنجش از دور ؛ جنگل تصادفی ; شاخص های پوشش گیاهی ; طبقه بندی یادگیری ماشین
1. مقدمه
علفزارهای دریایی در اکثر سواحل و مصب های دریایی کم عمق، سرپناه و با کف نرم در سرتاسر جهان وجود دارند و در میان مولدترین سیستم ها در اقیانوس قرار دارند [ 1 ، 2 ، 3 ]. آنها عملکردهای مهم تغذیه ای و ساختاری را در اکوسیستم های ساحلی انجام می دهند [ 4 ، 5 ، 6 ] و در بین زیستگاه های اولویت طبقه بندی شده توسط کنوانسیون تنوع زیستی (1992) قرار می گیرند. آنها به عنوان یکی از پنج عنصر کیفیت بیولوژیکی در ارزیابی کیفیت اکولوژیکی در آب های دریایی توسط دستورالعمل چارچوب آب اروپا (2000/60/EC) [ 7 ] فهرست شده اند. بنابراین، به دلیل کاهش گسترده بسترهای علف دریایی در مقیاس جهانی [1 ]، نظارت بر این اکوسیستم ها یک گام اساسی برای هر برنامه ای برای مدیریت این زیستگاه های کلیدی است. تکنیکهای زیادی برای پایش بسترهای علفدریایی استفاده میشود، با این حال بیشترین شاخصهای مورد استفاده در سراسر جهان به وسعت، تکه تکه شدن، محدودیتهای عمق سنجی (استعمار در عمق پایین)، فراوانی، نرخ بازیابی، زیست توده، تراکم، ارتفاع تاج و بیومتری برگ، ترکیب خاص و پوشش، جلبک های بزرگ مرتبط و بی مهرگان کفزی مرتبط [ 8 ].
نظارت بر وسعت علفزارهای دریایی یک رویکرد مفید برای تخمین اثرات عوامل استرس زای طبیعی و انسانی است. در واقع، این شاخص ساده ترین راه برای مطالعه این زیستگاه ها در مقیاس کوچک و بزرگ است [ 9 ]; کمترین تأثیر را بر محیط زیست دارد و میتواند نشانههایی از تخریب را نشان دهد و وجود اختلالات انسانی مانند اتروفیکاسیون، تغییر در استفاده از فضا و افزایش فعالیتهای آبی را نشان دهد. علاوه بر این، وجود یا عدم وجود بسترهای علف دریایی یا توزیع منطقه ای آنها محدودیت هایی را برای نمونه برداری از پوشش، زیست توده یا تراکم در امتداد شیب های عمقی مشخص می کند که شاخص های تکمیلی سلامت بسترهای علف دریایی هستند [ 8 ].
علف مارماهی کوتوله Z. noltei Hornemann، 1832، یکی از گونه های جهانی غالب در مناطق جزر و مدی است، بنابراین نمایانگر سطح مشترک زمین و دریا است [ 10 ]. پراکنش Z. noltei از سواحل اقیانوس اطلس شرقی از موریتانی تا جنوب نروژ و دریای کاتگات و در سرتاسر دریای مدیترانه، سیاه، آزوف، خزر و دریاهای آرال و جزایر قناری [ 11 ، 12 ، 13 ] گسترش می یابد. تختهای Z. noltei زیستگاهی را نشان میدهند که ساختار فضایی بسیاری از جوامع بیولوژیکی را امکانپذیر میسازد و اساس محفظههای تغذیهای استثنایی برای بسیاری از مصرفکنندگان اولیه در نظر گرفته میشود [ 14 ]. علاوه بر این،تختهای Z. noltei میتوانند به طور کمی به عملکرد آپارتمانهای جزر و مدی کمک کنند، زیرا آنها یک زیستگاه نوجوانان گسترده برای برخی از گونههای بسیار مهم در شبکه غذایی هستند [ 15 ]. در واقع، بووارور و همکاران. [ 16 ] نشان داد که زیست توده ماکروفونال رابطه مثبت معناداری با حضور علف مارماهی کوتوله دارد. آنها نشان دادند که تغییرات در ساختار جانوران macrobenthic عمدتا به دلیل وجود Z. noltei است. در مناطق ساحلی نیمه بسته، این گونه به ویژه در برابر اثرات ناشی از تغییرات آب و هوا و فشارهای انسانی آسیب پذیر است [ 17 ]. علیرغم اینکه Z.nolteiتوسط اتحادیه بین المللی حفاظت از طبیعت (IUCN) به عنوان گونه ای با نگرانی جزئی ذکر شده است، جمعیت کلی گزارش شده است که در حال کاهش است. Z. noltei علفزارهایی را عمدتاً در منطقه جزر و مدی تشکیل می دهد که احتمال آسیب پذیر بودن آن را به ویژه در برابر تأثیرات ناشی از تغییرات آب و هوایی، عمدتاً دمای آب دریا و افزایش افزایش سطح دریا (SLR) می کند، که بر وضعیت، ترکیب تأثیر می گذارد. و تکامل مناطق اشغالی، از بخش های همگن تا تکه های تکه تکه شده پراکنده در سواحل [ 14 ، 18 ، 19 ، 20 ].
مطالعات در طول زمان نشان داده است که کاربردهای سنجش از دور برای اکوسیستمهای علفدریایی ابزارهای خوبی برای نظارت هستند. این را می توان در سطح اولیه (تشخیص علف های دریایی، پوشش فضایی، تبعیض در سطح گونه ها، تشخیص زیست توده، الگوهای رشد و تخریب) یا در سطح ثانویه (متغیرهای محیطی موثر بر علف های دریایی، مانند دمای سطح دریا (SST) به دست آورد. شوری، افزایش سطح دریا، آلودگی، تشخیص اپی فیت ها و غیره) [ 21 ، 22 ، 23 ، 24 ، 25 ]. ترکیب تکنیک های سنجش از دور با بررسی های میدانی هنگام پایش وضعیت علف های دریایی و تغییرات در مقیاس های زمانی طولانی در مناطق بزرگ ضروری است [ 26 ، 27 ]، 28 ]. مراتع Z. noltei غالب ترین مناطق علف دریایی در سیستم های ساحلی نیمه محصور (SECS) سواحل اقیانوس اطلس مراکش هستند [ 29 ، 30 ]. جنبه های مربوط به نقشه برداری و نظارت طولانی مدت علف مارماهی کوتوله بسیار ناقص یا حتی وجود ندارد.
هدف مطالعه حاضر ترسیم چمنزارهای Z. noltei در تالاب ساحلی Merja Zerga با استفاده از تصویر SPOT 7 ارائهشده توسط مرکز سلطنتی سنجش از دور، همراه با دادههای میدانی بود. این اولین گام به سمت نظارت آینده و یک ابزار مهم برای مدیریت مراتع Z. noltei در امتداد سواحل اقیانوس اطلس مراکش است.
2. مواد و روشها
2.1. منطقه مطالعه
تالاب Merja Zerga که تالاب Moulay Bousselham نیز نامیده می شود، یک تالاب رامسر است. این تالاب شمالی ترین تالاب ساحلی در امتداد سواحل مراکش اقیانوس اطلس است و تقریباً در 120 کیلومتری شمال رباط، بین 34 درجه و 47 دقیقه و 34 درجه و 53 دقیقه شمالی و 6 درجه و 13 دقیقه و 6 درجه و 19 دقیقه غربی واقع شده است. این تالاب روزانه نفوذ جزر و مد اقیانوس اطلس را دریافت می کند که منبع اصلی حجم آب آن محسوب می شود. همچنین آب شیرین را از دو جریان اصلی دریافت می کند – کانال نادور که منشأ مصنوعی دارد و شامل آب زهکشی است و همچنین رودخانه درادر. علاوه بر این، تالاب دستخوش تغییرات انسانی قابل توجهی می شود [ 31 ]. در واقع، جمعیت مجاور این تالاب در 25 سال گذشته 150 درصد افزایش یافته است [ 32 ].
دمای آب این تالاب در تابستان بین 27 تا 28 درجه سانتیگراد و در زمستان از 13 تا 15 درجه سانتیگراد است [ 33 ]. عمق تالاب بین 0 تا 2 متر در نوسان است اما با توجه به چرخه جزر و مد و بارش به طور متوسط از 50 سانتی متر تجاوز نمی کند [ 34 ، 35 ]. جزر و مد دارای میانگین دامنه جزر و مدی نزدیک به 1.4 متر است که در طول جزر و مد بهار به 3.60 متر افزایش می یابد، در حالی که در طول جزر و مد به حدود 0.80 متر کاهش می یابد [ 36 ].
تالاب Merja Zerga از طریق بسیاری از فعالیت ها مانند گردشگری و شیلات، حمایت اجتماعی و اقتصادی مهمی را برای جوامع محلی فراهم می کند. علاوه بر این، منبع زیست محیطی قابل توجهی است که توجه بسیاری از محققان علمی را به خود جلب کرده است. این تالاب در طول دو دهه گذشته بخشی از پروژه های کاسارینا و ملمارینا بوده است [ 37 ]. بنابراین، مطالعات متعددی بر اساس مشاهدات و کار میدانی در Merja Zerga انجام شده است که عمدتاً ویژگی های هیدرولوژیکی، تغییرات محیطی، آلاینده های رسوبات، پلانکتون ها و جانوران ماهی را درمان می کند [ 34 ، 35 ، 38 ، 39 ، 40 ، 41 ، 42 ].
2.2. داده های میدانی
نمونههای علف دریایی از هفت ایستگاه علف مارماهی کوتوله در 4 تا 5 نوامبر 2019 گرفته شد و در 11 نوامبر 2019 به طور تصادفی بسته به شرایط جزر و مد انتخاب شدند و در تمام مناطق پوشش گیاهی آبی قابل تشخیص (گذر، شرق، غرب و مرکز شهر) پراکنده شدند. تالاب مرجا زرگا) از تحقیقات میدانی و مشاهدات تصاویر ماهواره ای ( جدول 1 ، شکل 1 ).
به منظور تعریف تفکیک پذیری طیفی تالاب گونه ها و مخلوط ها، اندازه گیری های طیفی در محل با استفاده از یک طیف سنج JAZA0478 با محدوده طول موج 348.95-1039.85 نانومتر، ارائه شده توسط آزمایشگاه LETG از دانشگاه نانت انجام شد. در هر ایستگاه، انعکاس انواع مختلف زیستگاه، به ویژه علف مارماهی کوتوله Z. noltei ، ماکرو جلبک (کلروفیت و رودوفیتا)، ماسه و رسوبات گل، در ارتفاع 1 متری از سطح زمین ثبت شد. برای هر نوع زیستگاه، به طور متوسط 3 تا 4 اندازه گیری طیفی در هر ایستگاه نمونه برداری شد و مراجع مربوطه آنها نیز اخذ شد. مراجع سفید و تیره با اندازه گیری بازتاب مرجع سفید ( Spectralon®WS-1-SL) و مرجع تیره (با پوشاندن لنز کولیمینگ با کلاهک آن انجام می شود).
به منظور تخمین زیست توده Z. noltei در گرم بر متر مربع و درصد پوشش آن، هفت ایستگاه در 16 سپتامبر 2018 در منطقه جزر و مدی واقع در پایین دست، بالادست و مرکز تالاب در هنگام جزر و مد نمونه برداری شد. زیست توده با استفاده از هسته های دستی (30 سانتی متر طول و 12 سانتی متر قطر)، با ده تکرار برای هر ایستگاه ارزیابی شد. نمونهها بهطور تصادفی از مراتع Z. noltei جمعآوری شدند، در محل شسته شدند و به آزمایشگاه منتقل شدند، سپس در دمای 20- درجه سانتیگراد قبل از اندازهگیری زیست توده منجمد شدند. نمونه ها قبل از توزین به مدت دو روز در فر در دمای 60 درجه سانتی گراد خشک شدند. برای تخمین درصد پوشش Z. noltei ، در هر ایستگاه، پنج تکرار کوادرات (0.5 × 0.5 متر مربع )) عکس گرفته شد و مختصات جغرافیایی آنها برای اعتبار سنجی نقشه ذخیره شد. عکسهای کوادراتها توسط نرمافزار ImageJ بر اساس پروتکلهای نظارت بر بستر ثابت برای دستورالعمل آب (WFD) Zostera marina (L.)- Z. noltei نسخه 3 [ 43 ] پردازش شدند. هنگامی که استفاده از کوادرات ممکن نبود، درصد پوشش به صورت بصری با اختصاص آن به یکی از کلاسهای زیر برآورد شد: 0-25٪، 25-50٪، 50-75٪، 75-90٪، 100٪.
2.3. تصاویر ماهواره ای
برای ترسیم توزیع علف مارماهی کوتوله Z. nolteiدر تالاب مرجا زرگا سعی کردیم تصاویر بسیار خوبی با وضوح بالا، در جزر و مد و با ضریب جزر و مد خوب به دست آوریم. این با تهیه جدول فهرستی با تاریخ جزر و مد، ضرایب و تعداد تصاویر حاوی ابر کمی به دست آمد. متأسفانه، نتایج بسیار محدود بود. باید بگوییم که این امر بر دشواری دستیابی به تصاویر بسیار خوب افزوده است. در نتیجه، ما از یک تصویر ماهوارهای SPOT 7 که در 19 اوت 2019 در ساعت 10:54 صبح به وقت محلی بهدستآمدیم، استفاده کردیم که با جزر و مد در حال سقوط مطابقت داشت. در واقع، 19 آگوست 2019 مربوط به کمترین جزر و مد است که 0.7 متر در ساعت 11:46 صبح UTC+1 با ضریب جزر و مد برابر با 72 بود. در اوت-سپتامبر به اوج خود رسید [ 44]. در واقع، در میان بسیاری از تصاویر Sentinel 2 و Pleiades، تصویر SPOT 7 بهترین شرایط مورد نیاز برای نقشه برداری از علف مارماهی کوتوله در تالاب Merja Zerga را ارائه می دهد.
ماهواره SPOT 7 دارای چهار باند چند طیفی آبی (450-520 نانومتر)، سبز (530-590 نانومتر)، قرمز (625-695 نانومتر) و مادون قرمز نزدیک (NIR؛ 760-890 نانومتر) – با وضوح فضایی است. 6×6 متر و یک نوار پانکروماتیک (450-745 نانومتر) با وضوح فضایی 1.5×1.5 متر [ 45 ]. صحنه عاری از پوشش ابری بود و توسط CRTS (مرکز سلطنتی سنجش از دور—مرکز سلطنتی تلویزیونی فضایی ( crts.gov.ma ، دسترسی به 25 فوریه 2021)) در قالب بستهای (PAN + XS) ارائه شد. که تصحیح شد و بر اساس داده های سیستم جهانی ژئودتیک 1984، کد EPSG: 4326 پیش بینی شد.
2.4. گونه شناسی
در رویکرد خود، زیستگاههای اصلی را که تالاب Merja Zerga را تشکیل میدهند با ارجاع به گونهشناسی فلورستیکی که توسط Hammada [ 46 ] برای این تالاب مشخص شده است، ترسیم کردیم. این گونه شناسی نشان داد که دو شیب وجود دارد که بر توزیع پوشش گیاهی تأثیر می گذارد، یعنی ریتم های غوطه وری و شوری. در واقع، گرادیان غوطه وری اجازه می دهد تا سه نوع گونه از هم متمایز شوند که عبارتند از:
-
گونه های مرطوب که تالاب ها را مستعمره می کنند (چمنزارهای مرطوب): Baldellia ranunculoides (L.) Parl.، Scirpus lacustris (L.) Palla.، Typha domigensis (Pers.) Steud.، Mentha aquatica (L.)، Nasturtium officinale (R. برادر)؛
-
گونه های آبزی، برخی از آنها در محیط های دریایی رشد می کنند ( Z. noltei ، Nitella sp. (CA) Agardh)، در حالی که بقیه در آب های آرام یا راکد مستعمره می شوند و می توانند شناور ( Lemna gibba (L.)) یا ثابت ( Myriophyllum sp. (L.), Potamogeton pectinatus (L.) Börner);
-
گونههای خشکیزی که در طول دوره بارانی در زیستگاههای لبهای معمولاً مرطوب رشد میکنند ( Centaurea calcitrapa (L.)، Euphorbia clementei Boiss.، Solanum nigrum (L.)).
هدف ما تشخیص گونه های آبزی محیط دریایی تالاب Merja Zerga بود. از این رو، طبقاتی که برای طبقه بندی مورد استفاده قرار گرفتند عبارتند از: آب کم عمق، ماسه، گل، گیاهان خشکی، جلبک، مخلوط جلبک و علف مارماهی کوتوله، علف مارماهی کوتوله غوطه ور و علف مارماهی کوتوله پدید آمده.
2.5. پردازش تصویر
تصویر SPOT 7 مورد استفاده برای این مطالعه در قالب بسته نرم افزاری (PAN + XS) تحویل داده شد. تصحیح اتمسفر برای چهار باند با استفاده از الگوریتم GRASS به منظور تبدیل آنها به بازتاب به دست آمد.
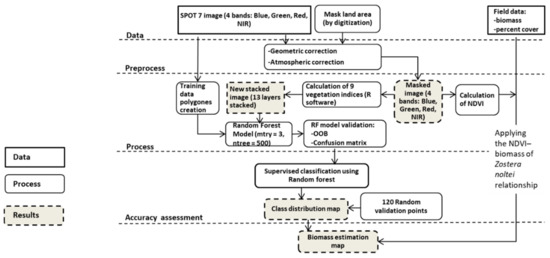
بنابراین، قدرت تفکیک طیفی و مکانی برای استفاده با 9 شاخص پوشش گیاهی مناسب بود ( جدول 1 ). در واقع، نقشه برداری از مارماهی کوتوله Z. noltei در تالاب Merja Zerga با استفاده از یک رویکرد طبقه بندی نظارت شده اعمال شده با الگوریتم جنگل تصادفی ( شکل 2 )، که مدلی است که تعداد زیادی از درختان تصمیم پایه را ساخته و ترکیب می کند، به دست آمد.
قبل از اعمال مدل جنگل تصادفی، 117 چند ضلعی به صورت دستی با استفاده از نرم افزار ArcGIS 10.4.1 بر اساس دانش میدان شخصی ایجاد شد و در هشت طبقه توزیع شد که برای طبقه بندی استفاده می شود: آب کم عمق، ماسه، گل، گیاهان خشکی، جلبک، مخلوطی از جلبک و علف مارماهی کوتوله، علف مارماهی کوتوله غوطه ور و علف مارماهی کوتوله پدید آمده. برای این کار، از جنگلهای تصادفی بریمن و کاتلر برای طبقهبندی و الگوریتم رگرسیون با بسته R randomforest v.4.6-14 استفاده کردیم. الگوریتم جنگل تصادفی که به عنوان یک مدل قوی و سازگار شناخته میشود، محبوبترین مدل مبتنی بر مجموعه برای مسائل طبقهبندی، و بهویژه برای نقشهبرداری از علفهای دریایی است [ 47 ].]. ما به طور تجربی مدل جنگلهای تصادفی را با آزمایش دو پارامتر تنظیم کردیم (زیرا به نظر میرسد که هیچ نظریهای برای هدایت انتخاب عدد mtry (تعداد پیشبینیکنندههای نمونهگیری شده برای تقسیم در هر گره) یا تعداد درختان وجود ندارد [ 48 ]. عدد mtry روی 3 ثابت شد (با استفاده از تابع tuneRF بسته تصادفی Forest) سپس با این عدد mtry و به منظور یافتن تعداد بهینه درختی که مطابق با یک طبقهبندی کننده پایدار بدون زمان محاسبات زیاد است. الگوریتم جنگلهای تصادفی را با مقادیر مختلف درخت محاسبه کرد و نرخ خطای OOB را برای ارزیابی تعداد درختهایی که نرخ خطای خارج از کیسه در آنها تثبیت شد و به حداقل رسید (در مورد ما 500 ثابت شد) ثبت کرد.
اول از همه، ما چهار باند خام تصویر SPOT 7 را برگرداندیم و از آنها برای محاسبه 9 شاخص پوشش گیاهی استفاده کردیم ( جدول 2 ). سپس لایههای بهدستآمده روی یک تصویر 13 باندی چیده شدند، که سپس با استفاده از نرمافزار R در مدل جنگلهای تصادفی پیادهسازی شد.
الگوریتم جنگلهای تصادفی با خطای مدل کلی خارج از کیسه (OOB) و ماتریس سردرگمی (درصد پیکسلهای آموزشی نادرست و طبقهبندیشده) تأیید شد و به نظر قوی بود (OOB = 3.14). سپس، ما از تابع پیش بینی برای محاسبه پیش بینی طبقه بندی RF با استفاده از رابط clusterR برای سرعت بخشیدن به محاسبات (در بسته R “راستر”) استفاده کردیم. علاوه بر این، تخمین نهایی کیفیت خطای طبقهبندی «دقت» با استفاده از نرمافزار Erdas با ایجاد 120 نقطه تصادفی مساوی شده در هشت کلاس انجام شد.
برای تخمین زیست توده بالای زمینی Z. noltei در بخش Merja Zerga از تالاب Moulay Bousselham با استفاده از سنجش از دور، ما انتخاب کردیم که از رابطه تجربی کمی NDVI – زیست توده Z. noltei استفاده کنیم که به موجب آن زیست توده = 610.61 (NDVI)^1.818 (n = ) r2 = 0.97)، همانطور که توسط Barillé و همکاران ایجاد شده است. (2010). بسته به وجود یا عدم حضور جلبک ها و شرایط ظهور یا غوطه وری، معادله پیشین برای درصد پوشش تخمینی جدا یا ترکیب شد ( شکل 2 ).
از این رو، یک نقشه تخمین زیست توده خاص برای هر یک از سه کلاس Z. noltei ایجاد شد: دو نقشه با اعمال مستقیم معادله برای هر دو کلاس Z. noltei غوطهور و کلاس Z. noltei ایجاد شد و یک نقشه ایجاد شد. با ترکیب معادله با درصد پوشش Z. noltei برای کلاس مخلوط جلبک و Z. noltei .
3. نتایج
3.1. نتایج طیفی
با توجه به طیف، تفکیک پذیری بین طبقات امکان پذیر بود. علاوه بر این، تصویر معادل کالیبره شده با بازتاب نیز تمایز بین طبقات را ممکن کرد. طیف بازتاب جلبک (n = 9) و Z. noltei (n = 13) نمونه برداری شده در تالاب Merja Zerga اشکال کلی متفاوتی را در محدوده مرئی و NIR بین 400 و 900 نانومتر ارائه کرد. در واقع، طیف جلبک در بالای طیف علف مارماهی کوتوله ترسیم شد. فاصله بین طیف جلبک ها و طیف های علف مارماهی کوتوله با استفاده از زاویه طیفی تخمین زده شد که دارای مقدار 0.936 رادیان بود. طیف بازتاب Z. nolteiاز کلروفیتا در طول موج های مرئی قابل تشخیص بودند. با این حال، آنها در NIR تمایل (افزایش) یکسانی داشتند، به جز طیف علف مارماهی کوتوله غوطه ور (کاهش) ( شکل 3 ). اشکال کلی طیف از تصویر SPOT 7 که ما استفاده کردیم مشابه آنهایی بود که با طیف رادیومتر در میدان نمونه برداری شد.
3.2. زیست توده و درصد پوشش Z. noltei
نتایج ما نشان داد که ایستگاه های S6، S4 و S7 دارای بالاترین زیست توده در بین پنج ایستگاه نمونه برداری شده در تالاب Merja Zerga بودند ( جدول 3 ).
با توجه به تفسیر بصری و تحلیل کوادرات، ایستگاه S7 کمترین درصد پوشش Z. noltei را داشت که به 50 درصد محدود شد ( جدول 4 ).
ایستگاه های S5 و S4 در جایگاه دوم قرار گرفتند که بسترهای مختلط ( Z. noltei و جلبک) با درصد پوشش Z. noltei به ترتیب به 60 و 75 درصد محدود شد. در مقابل، بقیه ایستگاه های تالاب بسترهای همگن با بیشترین درصد پوشش Z. noltei بودند که به 96 درصد رسید. در واقع، علیرغم اندازه کوچکشان، بسترهای علف مارماهی کوتوله در ایستگاه های “S3″، “S2” و “S1” همگن و متراکم تر بودند، به علاوه حضور گونه های ماکرو جلبک در آنجا چندان رایج نبود ( شکل 4 ).
3.3. پردازش تصویر ماهواره ای
مدل جنگلهای تصادفی ما توزیع هر هشت طبقه را برای تالاب Merja Zerga به دقت پیشبینی کرد. خطاهای کلاس داخلی مدل بین 0.00 تا 0.25 درصد در نوسان بود و OOB برای مدل 3.14 درصد بود. مشخص شد که باندها و شاخصهای خامی که برای تمایز بین هشت طبقه با توجه به میانگین کاهش شاخص جینی مرتبط بودند، شاخصهای NDVI و GNDVI و پس از آن GVI، CVI و EVI و سپس سبز و آبی هستند. باندهای متوالی ( شکل 5 ). در واقع، شاخصهای NDVI و GNDVI بالاترین میانگین کاهش مقادیر جینی را داشتند که به ترتیب به 03/141 و 69/138 رسید.
منحنی میانگین پیکسل ها در هر کلاس استخراج شده از تصویر SPOT 7 نشان داد که هر کلاس دارای باند یا شاخص خاصی است که به بهترین وجه آن را متمایز می کند. به عنوان مثال، شاخص IB برای تشخیص علف مارماهی کوتوله غوطه ور بیشترین ارتباط را داشت ( شکل 6 ).
3.4. طبقه بندی تصویر
ما موفق به برآورد مساحت مراتع Z. noltei در تالاب Merja Zerga شدیم که تقریباً برابر با 160.76 هکتار (شامل بسترهای مختلط) بود که از این مقدار 65.22 هکتار کل مساحت Z. noltei ظهور و غوطه ور بود .
نتایج طبقه بندی نشان داد که بسترهای Z. noltei در مرکز و در ناحیه بالادست تالاب نسبت به ناحیه پایین دست آن ( شکل 7 )، به ویژه در ایستگاه S6 ( جدول 5 ) گسترده تر است که با میدان همخوانی داشت. مشاهدات بنابراین، مجموع سطح برای بستر Z. noltei به ترتیب 27.31 هکتار، 95.54 هکتار، و 37.91 هکتار برای بستر مستغرق، مخلوط، و پدید آمده بود.
نتایج زیست توده علف مارماهی کوتوله با استفاده از محدوده سنجش از دور بین 0 تا 277.81 گرم DW/m2 برای کلاس ” Emerged Z. noltei ” و بین 0 تا 156.509 گرم DW/m2 برای کلاس “تخت های مختلط” تخمین زده شد که 62٪ آن (که میانگین سه ایستگاه مختلط است: “S5”: 0.77، “S4”: 0.60، “S7”: 0.50) با Z. noltei مطابقت داشت ، یعنی بین 0 تا 97.03 گرم DW/m2 بود . در نهایت، مقادیر زیست توده بین 0 تا 160.79 گرم DW/m2 برای کلاس ” Z. noltei غوطه ور ” بود ( شکل 8 ).
4. بحث
شکل کلی امضای طیفی جلبک ها و علف مارماهی کوتوله که در مزرعه یافت می شود، شبیه شکل های بدست آمده توسط Bargain و همکاران است. [ 54 ] و توسط Dehouck و همکاران. [ 57 ]. در واقع، بازتاب Z. noltei از جلبک های سبز در طول موج های مرئی و حتی بیشتر در طول موج NIR قابل تشخیص است. علاوه بر این، طیفهای علف مارماهی کوتوله و جلبک در طول موج NIR در مقایسه با طیفهای علف مارماهی کوتوله غوطهور افزایش مییابد که به دلیل وجود لایههای آب باقیمانده در آنها شیب کاهشی دارند. این به دلیل تأثیر آب است. Dehouck و همکاران [ 57 ] نشان داد که به طور کلی طیف های Z. nolteiو جلبک ها مشابه هستند، با این حال تفاوت های کوچک در نوارهای باریک قابل توجه است. آنها ادعا کردند که برای یافتن بهترین باندهای طیفی، طیف ها باید به صورت عددی تجزیه و تحلیل شوند.
ما انتخاب کردیم که از شاخصهای پوشش گیاهی برای نقشهبرداری Z. noltei در تالاب Merja Zerga استفاده کنیم، زیرا به شکل بسترهای علف دریایی تک گونهای وجود دارد. در واقع، منحنی میانگین پیکسلها در هر کلاس استخراجشده از تصویر SPOT 7 نشان داد که هر کلاس دارای باند یا شاخص خاصی است که به بهترین وجه آن را متمایز میکند. به عنوان مثال، شاخص IB برای تشخیص علف مارماهی کوتوله غوطه ور مرتبط ترین است ( شکل 6 ). نتایج جینی OOB و میانگین کاهش ما نشان داد که NDVI، GNDVI، GVI و CVI بهترین شاخصها برای استفاده برای این منظور هستند ( شکل 5 ). نقشه برداری از علفزارهای دریایی با استفاده از تصاویر چند طیفی و فراطیفی اغلب با استفاده از NDVI، که پرکاربردترین شاخص برای ارزیابی زیست توده سبز است، به دست آمده است [ 58 ]]. در واقع، Barillé و همکاران. [ 59 ] تصاویر SPOT 7 را به NDVI کالیبره کرد و تأثیر تغییرات زیست توده بالای زمینی Z. noltei را بر بازتاب طیفی آزمایش کرد. از این رو، آنها از داده های فراطیفی اندازه گیری شده میدانی از علف های دریایی و جلبک ها استفاده کردند و سپس از یک رابطه تجربی کمی NDVI-زیست توده برای ترسیم توزیع زیست توده Z. noltei استفاده کردند . دومی یک شیب تند مشخصه از 700 تا 900 نانومتر را نشان داد که با افزایش زیست توده افزایش یافت. بعداً، بارگین و همکاران. [ 60] نشان داد که شاخصهای پوشش گیاهی مورد استفاده برای نقشهبرداری این نوع تختها و زیست توده آنها با تصاویر ماهوارهای یا هوابرد باید با دقت انتخاب شوند تا همه منابع تغییرپذیری که بر مقدار شاخص تأثیر میگذارند، مانند تغییرات رنگدانه، یا منابع مربوط به تأثیرات پسزمینه، مانند کنتراست طیفی خاک، تغییرات albedo یا بازتاب دیدی در سطوح برگ.
مطالعه ما اولین مطالعه نقشه برداری از بسترهای Z. noltei با استفاده از الگوریتم جنگل تصادفی است. این روش نتایج رضایتبخشی را نشان داد زیرا برای تخمین پراکندگی سطحی این گونه با استفاده از تصاویر ماهوارهای SPOT 7 موثر ثابت شده است. نتایج نقشه برداری ما نشان داد که بسترهای Z. noltei در تالاب Merja Zerga گسترده تر از مطالعات قدیمی است [ 61 ، 62 ]. در واقع، مساحت سطح آنها به 65.22 هکتار رسید و با احتساب تشک های جلبک دریایی، این مقدار برابر با 160.76 هکتار بود ( جدول 6 ).
معمولاً نقشه برداری از مراتع Z. noltei با استفاده از ابزار GPS دستی یا با استفاده از ترانسکت زمانی که برخی از مناطق به دلیل ماهیت بستر، وسعت چمنزارها و چرخه های جزر و مدی غیر قابل دسترس هستند، کافی نیست. به طور کلی، تخمین توزیع علفهای دریایی با استفاده از روشهای سنتی با الگوهای شبکه از پیش ثبتشده یا ترکیبی از ترانسکتها و نقاط نمونهبرداری [ 63 ] زمانبر بوده و هزینههای مرتبط بالایی دارد [ 10 ]. بنابراین، می توان به لطف سنجش از راه دور، یا با تفسیر عکس های هوایی از یک وسیله نقلیه هوایی بدون سرنشین (UAV) همراه با اندازه گیری های حقیقت یابی زمینی [ 4 ، 5 ، 6 ، 7 ] به دست آورد.] یا با طبقه بندی عکس های زمینی، مایل، مقیاس بزرگ و تصاویر ماهواره ای [ 64 ، 65 ، 66 ، 67 ].
برخی از مطالعات نیز کارایی روش های تصادفی جنگل را در نقشه برداری از وسعت علف های دریایی ثابت کردند. در واقع، با توجه به Traganos و Reinartz [ 68 ]، طبقهبندیکنندههای تصادفی یادگیری ماشین جنگل و ماشینهای بردار پشتیبان علاقه بیشتری به سنجش از دور زیستگاه ساحلی پیدا میکنند. در واقع، آنها میزان تغییر Posidonia oceanica (L.) Delile و Cymodocea nodosa (Ucria) Ascherson را به دنبال تصحیح ستون آب اتمسفر و تحلیلی و با استفاده از یک الگوریتم جنگل تصادفی برای سری زمانی RapidEye ارزیابی کردند. علاوه بر این، ژانگ [ 69 ] و تراگانوس و راینارتز [ 70] ثابت کرد که الگوریتمهای جنگل تصادفی نتایج امیدوارکنندهای را در مورد طبقهبندی علفهای دریایی تولید میکنند، در حالی که Zhang و همکاران. [ 71 ] مزایای al را نشان داد. [ 57 ] تاکید می کند که با انتخاب دقیق باندهای چند طیفی یا با ترکیب آنها الگوریتم های جنگل تصادفی بر روی طبقه بندی کننده حداکثر احتمال با استفاده از تصاویر فراطیفی.
برخلاف مطالعه ما، چندین مطالعه قبلی وجود Z. noltei را در تالاب مرجا زرگا ذکر کرده اند، اما توزیع کلی آن را ذکر نکرده اند. ابتدا داکی و همکاران. [ 61 ] تخمین زد که 4.78 درصد از سطح تالاب را مراتع علف مارماهی کوتوله اشغال کرده است که مساحت سطح آن با 140 هکتار از جمله گونه های جلبک مطابقت دارد. وجود Z. noltei در تالاب مرجا زرگا نیز توسط بوتاهر و همکاران گزارش شده است. [ 30 ]، گام و همکاران. [ 72 ]، Grignon-Dubois و Rezzonico [ 73 ]، Natij و همکاران. [ 74 ]، توهامی و همکاران. [ 75 ] و توهامی و همکاران. [ 76]. علفزارهای کوتوله علف مارماهی در این تالاب در بسترهای گل آلود و گاهی عمیق ایجاد شده اند که راه رفتن از میان آنها را بسیار دشوار می کند، بنابراین نقشه برداری از این علفزارها با پای پیاده امکان پذیر نیست. اخیراً به گفته زرانز السو و همکاران. [ 62 ]، علف مارماهی کوتوله در تالاب Merja Zerga از طریق خصوصیات طیفی و طبقهبندی حداکثر احتمال تصویر ماهوارهای World View 2 که در 3 نوامبر 2010 به دست آمد، همراه با نمونههای میدانی نقشهبرداری شده است. از این رو، مساحت مراتع Z. noltei در تالاب مرجا زرگا 71.508 متر مربع برآورد شد .با این حال در این مورد تصویر ماهواره ای استفاده شده تمام مناطق جزر و مدی تالاب را پوشش نمی دهد. با این حال، این مطالعه بسترهای بزرگتر واقع در منطقه پایین دست تالاب را از دست داد. هنگام تفسیر تصاویر نوری چندطیفی، سردرگمی بین علفزارهای کوتوله مارماهی، جلبکها و علف طناب Spartina sp.، که یک گونه نوظهور هالوفیل است که در تالاب با سایر گونههای شورهزار، بهویژه Salicornia sp، هم مرز است، ادامه داشت. (L.) و Juncaceae راش [ 62 ، 75 ]. Dehouck و با استفاده از یک پایگاه داده چند زمانی توانستند این سردرگمی را محدود کنند و به نظر می رسد که جلبک ها فقط در هنگام استفاده از باندهای طیفی خوب قابل تشخیص هستند. به گفته Proença و همکاران. [ 77 ]، نقشه برداری ازSpartina anglica CE Hubb. و Spartina maritima (Curtis) Fernald با استفاده از طبقهبندی نظارت شده تصاویر Pleiades نشان دادند که این گونهها اغلب با مناطق وسیعی که توسط Z. noltei پوشش گیاهی دارند ، بهویژه در ماه اکتبر اشتباه گرفته میشوند. برای دستیابی به نتایج بهتر و جلوگیری از این سردرگمی، بهتر است قبل از اجرای الگوریتم طبقهبندی، اکثر پوشش گیاهی بالقوه زمین را از تصویر ماهوارهای پوشانده شود. در مورد ما، این مشکل رخ نداد زیرا ما تعداد کافی چند ضلعی (117) ایجاد کردیم که در بین هشت کلاس مورد استفاده برای طبقهبندی توزیع شدهاند. در واقع، بر اساس نتایج ارزیابی دقت، طبقهبندی تصادفی جنگل یک Z. noltei رضایتبخش ایجاد کرد.نقشه چمنزار، با شاخص کاپا برابر با 0.9429 و دقت کلی برابر با 95٪.
نتایج طبقهبندی ما امکان تخمین وسعت کلی Z. noltei در تالاب Merja Zerga را فراهم کرد و همچنین امکان تخمین زیست توده بالای زمینی این گونه را فراهم کرد. همانطور که با مطالعه Barillé و همکاران. [ 60 ]، در مورد ما، رابطه کمی NDVI-زیست توده برای همه مراتع Z. noltei با موفقیت اعمال شد، زیرا با وجود انباشته شدن بسیاری از لایه ها در مناطق خاص، NDVI اشباع نشده بود. بنابراین، این روش مستقیماً برای کلاسهای « Emerged Z. noltei » و «Submerged Z. noltei » به کار گرفته شد و با درصد پوشش برای کلاس «مخلوط Z. noltei و جلبک» ترکیب شد. در واقع، ما در طول تحقیقات میدانی مشاهده کردیم کهZ. noltei ماکروفیت اصلی زنده در منطقه جزر و مدی نقشهبرداری شده نبود – ماکروجلبکهای سبز رشتهای در بسترهای مختلط مشاهده شدند و چند گونه ماکروجلبک (Cholorophyta و Rhodophyta) گاهی اوقات وجود داشتند ( شکل 9 ).
در واقع، واله و همکاران. [ 78 ] ادعا کرد که NDVI برای تمایز بین بسترهای با پوشش گیاهی کم و متراکم Z. noltei مفید است ، و بنابراین میتواند بیشتر در تصاویر CASI (حسگر فراطیفی) برای نقشهبرداری درصد پوشش گونههای ساکن در منطقه اعمال شود.
کالیبراسیون تصویر SPOT 7 با رابطه زیست توده-NDVI الگویی از حداکثر زیست توده را در طبقات غوطه ور و ظهور نشان داد، به ویژه در ایستگاه “S6″، و زیست توده پایین تر در منطقه مرکزی (ایستگاه های “S5” و “S4” ). میانگین زیست توده اندازه گیری شده در محل در سپتامبر 2018 تفاوت معنی داری با مقدار به دست آمده از سنجش از دور نداشت ( جدول 7 ).
اخیراً، دادههای سنجش از راه دور فعال از هواپیماها یا پهپادها برای نقشهبرداری از پوشش علفهای دریایی، عمدتاً LiDAR استفاده شده است [ 78 ، 79 ، 80 ، 81 ، 82 ، 83 ، 84 ، 85 ، 86 ]. حسگرهای فراطیفی مانند CASI-2، PHILLS و HyMap [ 78 ، 87 ، 88 ، 89 ، 90 ]؛ یا عکسهای [ 91 ، 92 ، 93 ، 94 ، 95]. کارایی داده های سنجش از دور از طریق بسیاری از مطالعات مربوط به تخمین پوشش علف های دریایی ثابت شده است. با این وجود، اکثر مطالعات هنوز به اندازه گیری های میدانی در هنگام تخمین تراکم ساقه و ارتفاع تاج متکی هستند (به عنوان مثال، [ 96 ]).
5. نتیجه گیری ها
در مطالعه ما، مناسب بودن استفاده از سنجش از دور برای نقشه برداری از توزیع Z. noltei و در مرحله دوم برای تخمین زیست توده آن تایید شد. در واقع، ما یک تصویر ماهوارهای SPOT 7 را با استفاده از الگوریتم جنگل تصادفی و دادههای میدانی برای تهیه نقشههای زیستگاه علف مارماهی کوتوله طبقهبندی کردیم. این رویکرد ما را قادر ساخت تا توزیع بسترهای Z. noltei و جلبک های بزرگ مرتبط را از نظر پوشش سطحی تعریف کنیم. نتایج ما با دقت 95 درصد ثابت کرد که GNDVI، NDVI، و EVI شاخصهای گیاهی بالقوه مفیدی برای تشخیص بسترهای علف مارماهی کوتوله از سایر زیستگاههای تالاب هستند. نقشههای قدیمیتر شامل توصیف فضایی Z. nolteiدر تالاب مرجا زرگا نمی توان وسعت کامل این علف دریایی را تعیین کرد. در مقایسه با کار ما، مطالعات قبلی روی Z. nolteiنقشه برداری وسعت آن را دست کم گرفت زیرا آنها از تصاویری استفاده کردند که با همه تالاب Merja Zerga مطابقت نداشت یا در جزر و مد بالاتر گرفته شده بود. در واقع، سطح کلی اشغال شده توسط علف مارماهی کوتوله در این تالاب با در نظر گرفتن بسترهای مختلط، 160.76 هکتار است. رویکرد ما بر اساس تخمین گستره است، که یک شاخص جهانی است که میتواند افزایش یا از دست دادن مساحت بسترهای علفهای دریایی را در محدوده آن آشکار کند. تلفات داخل این محدوده ها را که در سطح تکه تکه شدن این بسترها قابل مشاهده است، در نظر نمی گیرد. تخمین زیست توده علف مارماهی کوتوله با استفاده از مدل کمی NDVI-biomass اعمال شده در تصاویر SPOT 7 می تواند به عنوان یک روش غیر مخرب برای تخمین زیست توده این گونه علف دریایی بسیار مفید باشد. در مطالعات آتی Z. nolteiو برای خرید داده های به دست آمده باید همین روش در نظر گرفته شود.
منابع
- Orth، RJ; Carruthers، TJ; دنیسون، WC; دوارته، سی ام. Fourqurean، JW; هک، KL; هیوز، آر. کندریک، GA; Kenworthy، WJ; اولیارنیک، S. یک بحران جهانی برای اکوسیستم های علف دریایی. Bioscience 2006 ، 56 ، 987-996. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- دوارته، سی. بوروم، جی. کوتاه، FT; واکر، اکوسیستمهای سیگرس DI: وضعیت جهانی و چشمانداز آنها. در اکوسیستم های آبی: روندها و چشم اندازهای جهانی ؛ انتشارات دانشگاه کمبریج: کمبریج، انگلستان، 2008; ص 281-294. [ Google Scholar ] [ CrossRef ]
- کوتاه، FT; پولیدورو، بی. لیوینگستون، SR; نجار، KE; باندیرا، اس. Bujang، JS; Calumpong، HP; Carruthers، TJB; کولز، آر جی. دنیسون، WC; و همکاران ارزیابی خطر انقراض گونه های علف دریایی جهان Biol. حفظ کنید. 2011 ، 144 ، 1961-1971. [ Google Scholar ] [ CrossRef ]
- دافی، جی. تنوع زیستی و عملکرد اکوسیستم های علف دریایی. مارس اکل. Prog. سر. 2006 ، 311 ، 233-250. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- کوتاه، اف. کاروترز، تی. دنیسون، دبلیو. Waycott، M. توزیع و تنوع علف دریایی جهانی: یک مدل زیست منطقه ای. J. Exp. مار. بیول. Ecol. 2007 ، 350 ، 3-20. [ Google Scholar ] [ CrossRef ]
- گوتیرز، جی.ال. جونز، سی جی; Byers, JE; آرکما، ک. برکنبوش، ک. Commito، JA; دوارته، سی ام. هکر، SD; Lambrinos، JG; هندریکس، IE; و همکاران مهندسان اکوسیستم فیزیکی و عملکرد مصب ها و سواحل. در رساله در علم رودخانه و سواحل ; انتشارات دانشگاهی: Waltham, MA, USA, 2011; جلد 7، ص 53-81. [ Google Scholar ]
- ماربا، ن. کراوس-جنسن، دی. آلکورو، تی. بیرک، اس. پدرسن، آ. نتو، جی.ام. اورفانیدیس، س. Garmendia, JM; موکسیکا، آی. بورجا، ا. و همکاران تنوع شاخص های علف دریایی اروپا: الگوهای درون و بین مناطق Hydrobiologia 2012 ، 704 ، 265-278. [ Google Scholar ] [ CrossRef ]
- فونتان، ای. دوما، پ. Ponton، D. Méthodes de Cartographie، de Caractérisation et de Suivi des Herbiers Marins. 2011. در دسترس به صورت آنلاین: https://www.google.com/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&cad=rja&uact=8&ved=2ahUKEwjenJLGiLHwAhWJFMAKHcgtA4gQFjAAegQIASyFestem% %2Ffiles_force%2Fdocuments%2Fmethode_de_cartographie_de_caracterisation_et_de_suivi_des_herbiers_marins.pdf%3Fدانلود%3D1&usg=AOvVaw3mGzT8NjBkgJVlHJK_Yj1Rac5 فوریه.
- بوروم، جی. دوارته، سی ام. کراوس-جنسن، دی. Greve, TM European Seagrasses: مقدمه ای بر نظارت و مدیریت . نظارت و مدیریت پروژه اتحادیه اروپا علفهای دریایی اروپا (پروژه M&MS): Hilleroed, Danmark, 2004; شابک 87-89143-21-3. [ Google Scholar ]
- کوتاه، FT; کولز، روش تحقیق جهانی علف دریایی RG ; Elsevier Science BV: آمستردام، هلند، 2001; 473p. [ Google Scholar ]
- گرین، ER; کوتاه، FT World Atlas of Seagrasses ; انتشارات دانشگاه کالیفرنیا: برکلی، کالیفرنیا، ایالات متحده آمریکا، 2003. [ Google Scholar ]
- مور، کالیفرنیا؛ کوتاه، FT Zostera: زیست شناسی، بوم شناسی و مدیریت. در Seagrasses: Biology, Ecology and Conservation ; Larkum, T., Orth, R., Duarte, C., Eds.; Springer: Dordrecht، هلند، 2006; صص 361-386. [ Google Scholar ]
- Diekmann، OE; گوویا، ال. پرز، جی. گیل رودریگز، سی. Serrão، EA منشاء احتمالی Zostera noltii در جزایر قناری و دستورالعملهایی برای بازسازی. مار. بیول. 2010 ، 157 ، 2109-2115. [ Google Scholar ] [ CrossRef ]
- Dalloyau، S. Qualité Ecologique des Herbiers Intertidaux à Zostère Naine Zostera Noltei dans les Sites Fonctionnels des Réserves Naturelles Nationales Gérées par la LPO (Pertuis Charentais). 2020. در دسترس آنلاین: https://www.researchgate.net/publication/349345847_Qualite_ecologique_des_herbiers_intertidaux_a_Zostere_naine_Zostera_noltei_dans_les_sites_fonctionnels_des_lles_Reserves_Noltei_dans_les_sites_fonctionnels_des_lles_Reserves_National .
- پولته، پی. شانز، آ. Asmus، H. سهم بسترهای علف دریایی ( Zostera noltii ) در عملکرد مناطق جزر و مدی به عنوان یک زیستگاه جوان برای epibenthos غالب و متحرک در دریای Wadden. مار. بیول. 2005 ، 147 ، 813-822. [ Google Scholar ] [ CrossRef ]
- بووارور، او. ال کامچا، ر. بوتومیت، اس. پوزت، پ. معنان، م. بزیری، H. اثرات مراتع Zostera noltei بر جانوران اعماق دریا در اکوسیستم های ساحلی اقیانوس اطلس شمالی مراکش: الگوهای فضایی و فصلی. Biologia 2021 ، 1-13. [ Google Scholar ] [ CrossRef ]
- کاباچو، اس. سانتوس، آر. Sprung, M. پویایی جمعیت و تولید علف دریایی Zostera noltii در استعمار در مقابل مراتع مستقر. مارس اکل. 2011 ، 33 ، 280-289. [ Google Scholar ] [ CrossRef ]
- چست، جی. بورجا، Á. کابالرو، آ. ایریگوین، ایکس. سانز، جی. مونچو، آر. مارکوس، ام. لیریا، پ. هیدالگو، جی. واله، ام. و همکاران تأثیرات تغییر آب و هوا بر محیط های ساحلی و دریایی در جنوب شرقی خلیج بیسکای. صعود Res. 2011 ، 48 ، 307-332. [ Google Scholar ] [ CrossRef ]
- ماسا، SI; Arnaud-Haond، S. پیرسون، GA; Serrão، EA تحمل دما و بقای جمعیت های جزر و مدی علف دریایی Zostera noltii (Hornemann) در جنوب اروپا (ریا فورموسا، پرتغال). هیدروبیول. 2008 ، 619 ، 195-201. [ Google Scholar ] [ CrossRef ]
- کوتاه، FT; Neckles, HA اثرات تغییرات آب و هوایی جهانی بر علفهای دریایی. آکوات. ربات. 1999 ، 63 ، 169-196. [ Google Scholar ] [ CrossRef ]
- شوو، اس. بوچون، سی. تکنیکهای سنجش از دور Maniere، R. سازگار با نقشهبرداری با وضوح بالا از اکوسیستمهای دریایی استوایی ساحلی (صخرههای مرجانی، بستر علفهای دریایی و حرا). بین المللی J. Remote Sens. 1998 , 19 , 3625-3639. [ Google Scholar ] [ CrossRef ]
- دکر، ا. براندو، وی. آنتسی، جی. فایف، اس. مالتوس، تی. کارپوزلی، الف. سنجش از دور اکوسیستمهای علف دریایی: استفاده از حسگرهای فضابرد و هوابرد. در Seagrasses: Biology, Ecology and Conservation ; Larkum, T., Orth, RJ, Duarte, CM, Eds. Springer: Dordrecht، هلند، 2006; صص 347-359. شابک 978-1-4020-2983-7. [ Google Scholar ]
- Ferwerda، JG; دی لیو، جی. آتزبرگر، سی. Vekerdy، Z. نظارت مبتنی بر ماهواره بر پوشش گیاهی علف های دریایی گرمسیری: تکنیک های فعلی و پیشرفت های آینده. Hydrobiologia 2007 ، 591 ، 59-71. [ Google Scholar ] [ CrossRef ]
- حسین، ام اس; Bujang، JS; زکریا، م.ح. هاشم، م. کاربرد تصاویر لندست در تحلیل تغییر پوشش منطقه علف دریایی برای لاواس، ترنگانو و کلانتان مالزی. ادامه Shelf Res. 2015 ، 110 ، 124-148. [ Google Scholar ] [ CrossRef ]
- حسین، ام اس; هاشم، ام. فناوریهای بالقوه رصد زمین (EO) برای ارزیابی خدمات اکوسیستم علفدریایی. بین المللی J. Appl. زمین Obs. Geoinf. 2019 ، 77 ، 15-29. [ Google Scholar ] [ CrossRef ]
- دینگتیان، ی. Chaoyu، YDAY توزیع علف دریایی در چین با سنجش از دور ماهواره ای. در سنجش از دور سیاره زمین ; IntechOpen: لندن، بریتانیا، 2012؛ پ. 29043. موجود آنلاین: https://www.intechopen.com/books/remote-sensing-of-planet-earth/seagrass-distribution-in-china-with-remote-sensin (در 25 فوریه 2021 قابل دسترسی است).
- مایر، کالیفرنیا؛ ارزیابی منابع Pu، R. Seagrass با استفاده از روشهای سنجش از دور در سنت جوزف ساوند و بندر کلیرواتر، فلوریدا، ایالات متحده آمریکا. محیط زیست نظارت کنید. ارزیابی کنید. 2011 ، 184 ، 1131-1143. [ Google Scholar ] [ CrossRef ]
- رولفسما، سی ام. Phinn, SR; Udy، NS; Maxwell، PS یک رویکرد میدانی و سنجش از دور یکپارچه برای نقشه برداری Seagrass Cover، Moreton Bay، استرالیا. جی. اسپات. علمی 2009 ، 54 ، 45-62. [ Google Scholar ] [ CrossRef ]
- بووارور، او. ال کامچا، ر. بوطاهر، ل. تنومی، ع. زوراره، بی. بنهوسا، ا. بازاری، H. الگوهای فضایی مراتع Zostera noltei در سراسر سواحل اقیانوس اطلس مراکش: آیا شیب عرضی وجود دارد؟ PeerJ PrePr. 2015 ، 3 ، e1076v1. [ Google Scholar ] [ CrossRef ]
- بوطاهر، ل. معنان، م. بووارور، او. ریچر، جی. پوزت، پ. گوبرت، اس. معنان، م. زوراره، بی. بنهوسا، ا. بذری، ح. پایش زیست محیطی در اکوسیستم های نیمه محصور ساحلی با استفاده از مراتع Zostera noltei. Ecol. اندیک. 2019 ، 104 ، 776-793. [ Google Scholar ] [ CrossRef ]
- احمد، م.ح. ال لیتی، BM; تامپسون، جی آر. گل، RJ; رمدانی، م. ایاچه، اف. حسن، اس ام کاربرد سنجش از دور در توصیف مکان و تحلیل تغییرات محیطی تالاب های ساحلی شمال آفریقا. Hydrobiologia 2009 ، 622 ، 147-171. [ Google Scholar ] [ CrossRef ]
- معنان، م. لندزمن، سی. معنان، م. زوراره، بی. فتال، پ. سحابی، م. ارزیابی هجوم انسانی آلاینده های فلزی و متالوئیدی به تالاب مولای بوسلهام، مراکش، با استفاده از روش های کمومتری همراه با سیستم های اطلاعات جغرافیایی. محیط زیست علمی آلودگی Res. 2013 ، 20 ، 4729-4741. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- بازاری، ح. باید، ا. گلمارک، ام. Hily، C. سازماندهی فضایی جوامع macrozoobenthic در پاسخ به عوامل محیطی در یک تالاب ساحلی از ساحل شمال غربی آفریقا (Merja Zerga، مراکش). اوشنول. Acta 2003 ، 26 ، 457-471. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- کرایم، م. بن حمزه، ج. رمدانی، م. فتحی، ع. عبدالظاهر، ح. Flower, R. برخی از مشاهدات در مورد سن و رشد کفال خاکستری لب نازک، Liza ramada Risso، 1826 (ماهی، Mugilidae) در سه دریاچه تالاب شمال آفریقا: Merja Zerga (مراکش)، Garâat Ichkeul (تونس) و دریاچه Edku ( مصر). آکوات. Ecol. 2001 ، 35 ، 335-345. [ Google Scholar ] [ CrossRef ]
- لابردی، ح. اطهیری، ا. لازار، اس. ماسیک، ز. El Antri، S. Étude de la variation spatio-temporelle des physico-chimiques caractérisant la qualité des eaux d’une lagune cotière et ses zoneions écologiques: Cas de Moulay Bousselham، Maroc. CR GEOSCI 2005 ، 337 ، 505-514. [ Google Scholar ] [ CrossRef ]
- علاوی، ع.م. چورا، م. معنان، م. زوراره، بی. رابین، ام. Conceição، MF; آندراد، سی. خالد، م. Carruesco، C. شار فلز به رسوبات تالاب Moulay Bousselham، مراکش. محیط زیست علوم زمین 2009 ، 61 ، 275-286. [ Google Scholar ] [ CrossRef ]
- گل، RJ; تامپسون، JR مروری بر مطالعات یکپارچه آبی-اکولوژیکی در پروژه MELMARINA: پایش و مدلسازی تالابهای ساحلی – ساخت ابزارهای مدیریتی برای منابع آبی در شمال آفریقا. Hydrobiologia 2008 ، 622 ، 3-14. [ Google Scholar ] [ CrossRef ]
- Appleby، PG; Birks، HH; گل، RJ; رز، ن. Peglar، SM; رمدانی، م. Kraïem، MM; فتحی، AA تاریخ ها و نرخ های رسوب گذاری رسوبات اخیر در نه دریاچه تالاب شمال آفریقا (پروژه کاسارینا) را به صورت رادیومتری تعیین کرد. آکوات. Ecol. 2001 ، 35 ، 347-367. [ Google Scholar ] [ CrossRef ]
- پیترز، آ. جونز، KC; گل، ر. اپلبی، پی. رمدانی، م. کرایم، م. فتحی، الف. تغییرات محیطی اخیر در دریاچههای تالاب آفریقای شمالی: مطالعه پایه باقی ماندههای آلاینده آلی کلر در رسوبات نه سایت در پروژه کاسارینا. آکوات. Ecol. 2001 ، 35 ، 449-459. [ Google Scholar ] [ CrossRef ]
- رمدانی، م. گل، RJ; الخیاتی، ن. Kraïem، MM; فتحی، ع.ا. Birks، HH; دریاچههای تالاب پاتریک، ST آفریقای شمالی: ویژگیهای نه سایت موجود در پروژه کاسارینا. آکوات. Ecol. 2001 ، 35 ، 281-302. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- رمدانی، م. الخیاتی، ن. گل، RJ; تامپسون، جی آر. چوبا، ال. Kraiem، MM; ایاچه، اف. احمد، MH تأثیرات محیطی بر ترکیب کیفی و کمی فیتوپلانکتون و زئوپلانکتون در دریاچه های ساحلی شمال آفریقا. Hydrobiologia 2009 ، 622 ، 113-131. [ Google Scholar ] [ CrossRef ]
- تامپسون، جی آر. گل، RJ; رمدانی، م. ایاچه، اف. احمد، م.ح. راسموسن، EK; پترسن، ویژگی های هیدرولوژیکی سیستم عامل سه تالاب ساحلی شمال آفریقا: بینش هایی از پروژه MELMARINA. Hydrobiologia 2009 ، 622 ، 45-84. [ Google Scholar ] [ CrossRef ]
- هیلی، سی. Sauriau، PG; Auby , I. Protocoles Suivi Stationnel des Herbiers à Zostères pour la Directive Cadre sur l’Eau (DCE) Zostera marina ; Rapport Ifremer/ODDE/UL/LER/AR/18.017؛ Ifremer: Brest، فرانسه، 2007. [ Google Scholar ]
- وانگ، ام سی؛ براوو، MA; داود، ام. پویایی زیست محیطی Zostera marina (علف مارماهی) در سه خلیج مجاور در اقیانوس اطلس کانادا. ربات. مارس 2013 ، 56 ، 413-424. [ Google Scholar ] [ CrossRef ]
- خدمات آستریوم راهنمای کاربر تصاویر SPOT 6 و SPOT 7. فرانسه. 2013. موجود به صورت آنلاین: https://www.google.com/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&ved=2ahUKEwj-r6nx_LDwAhWC7eAKHQ8lBF8QFjABegQIAhAD&url=2Face%Office.Fb . %2FUser_Guide_SPOT6_V1.0.pdf&usg=AOvVaw1ppInVLdBsoFQgGFiigs5a (دسترسی در 25 فوریه 2021).
- Hammada، S. Etudes sur la Vegétation des zones Humides du Maroc: Catalog et Analyze de la Biodiversité Floristique et Identification des Principaux Groupements Végétaux، 2017 ; Université Mohammed V-Agdal, Faculté des Sciences Rabat: Rabat, Morocco, 27 فوریه 2007. [ Google Scholar ]
- ها، NT; مانلی هریس، ام. فام، تی دی; هاوز، I. ارزیابی مقایسه ای روش های یادگیری ماشینی مبتنی بر گروه و حداکثر احتمال برای نقشه برداری علف دریایی با استفاده از تصاویر Sentinel-2 در بندر تاورانگا، نیوزیلند. Remote Sens. 2020 , 12 , 355. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- Scornet، E. پارامترهای تنظیم در جنگل های تصادفی. ESAIM: Proc. Surv. 2017 ، 60 ، 144-162. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- Gitelson، AA; کافمن، YJ; Merzlyak، MN استفاده از یک کانال سبز در سنجش از دور پوشش گیاهی جهانی از EOS-MODIS. سنسور از راه دور محیط. 1996 ، 58 ، 289-298. [ Google Scholar ] [ CrossRef ]
- Rouse, JWJ; هاس، RH; شل، JA; Deering، DW نظارت بر سیستم های پوشش گیاهی در دشت های بزرگ با ERTS. مشخصات ناسا انتشار 1974 ، 351 ، 309-317. [ Google Scholar ]
- Kauth، RJ; توماس، GS کلاهک منگولهای – یک توصیف گرافیکی از توسعه طیفی-زمانی محصولات کشاورزی همانطور که توسط Landsat دیده میشود. در سمپوزیوم LARS ; دانشگاه پردو: وست لافایت، IN، ایالات متحده آمریکا، 1976. [ Google Scholar ]
- اسکادافل، ر. Huete, A. بهبود سنجش از دور پوشش گیاهی کم در مناطق خشک با اصلاح شاخص های پوشش گیاهی برای “صدای” خاک. Etude des propriétés spectrales des sols arides appliquée à l’Alélioration des indices de vegétation obtenus par télédétection. فائو CR Acad. علمی 1991 ، 312 ، 1385-1391. [ Google Scholar ]
- گوئل، NS; Qin، W. تأثیرات معماری تاج پوشش بر روابط بین شاخصهای مختلف پوشش گیاهی و LAI و Fpar: یک شبیهسازی کامپیوتری. Remote Sens. Rev. 1994 , 10 , 309-347. [ Google Scholar ] [ CrossRef ]
- هیوت، ا. عدالت، سی. لیو، H. توسعه پوشش گیاهی و شاخص های خاک برای MODIS-EOS. سنسور از راه دور محیط. 1994 ، 49 ، 224-234. [ Google Scholar ] [ CrossRef ]
- Rapinel, S. Contribution de la Télédétection à l’évaluation des Fonctions des Zones Humides: De l’Observation à la Modélisation Prospective ; تاریخچه؛ Université Rennes 2: Rennes، فرانسه، 2012. [ Google Scholar ]
- هیوت، ا. جکسون، آر. خاک و جو بر روی طیف تاج پوشش های جزئی تأثیر می گذارد. سنسور از راه دور محیط. 1988 ، 25 ، 89-105. [ Google Scholar ] [ CrossRef ]
- Dehouck، A. لافون، وی. لوباک، بی. کرولا، اس. برو، دی. اشملتز، ام. Roubache، A. پایگاه داده میدان ابرطیفی در پشتیبانی از نقشه برداری تالاب ساحلی. در مجموعه مقالات سمپوزیوم بین المللی زمین شناسی و سنجش از دور IEEE 2012، مونیخ، آلمان، 22 تا 27 ژوئیه 2012. صص 2649–2652. [ Google Scholar ] [ CrossRef ]
- لیلسند، MT; Kiefer, R. Remote Sensing and Image Interpretation , 4th ed.; جان وایلی و پسران: نیویورک، نیویورک، ایالات متحده آمریکا، 2000; 736p. [ Google Scholar ]
- باریله، ال. رابین، ام. هارین، ن. چانه زنی، ا. Launeau، P. افزایش توزیع علف دریایی در خلیج Bourgneuf (فرانسه) توسط سنجش از دور فضایی شناسایی شد. آکوات. ربات. 2010 ، 92 ، 185-194. [ Google Scholar ] [ CrossRef ]
- چانه زنی، ا. رابین، ام. ملدر، وی. رزا، پ. لو من، ای. هارین، ن. باریل، L. تغییرات طیفی فصلی Zostera noltii و تأثیر آن بر شاخصهای گیاهی مبتنی بر رنگدانه. J. Exp. مار. بیول. Ecol. 2013 ، 446 ، 86-94. [ Google Scholar ] [ CrossRef ]
- داکی، م. بنهوسا، ا. Hammada، S. ابن تطو، م. قنینبا، ع. El Agbani، MA Cartographie des Habitats Naturels et de la Vegétation de Merja Zerga، Maroc. رپ ؛ Inédit, MedWet 2; Bureau Ramsar/Administration des Eaux & Forêts et de la Conservation du sol: داکار، سنگال، 1998. [ Google Scholar ]
- Elso، MZ; ماننت، پی. لوک، ا. رمدانی، م. Robaina، R. توصیف ژنتیکی و تکنیکهای سنجش از دور به عنوان ابزارهای مدیریتی برای جمعیتهای علف دریایی Zostera noltii در امتداد سواحل مراکش اقیانوس اطلس. جی. ساحل. Res. 2017 ، 33 ، 78-87. [ Google Scholar ] [ CrossRef ]
- قنینبا، ع. بنهوسا، ا. الاقبانی، م.-ع. داکی، م. Thevenot, M. Etude phénologique et variabilité interannuelle d’abondance des Charadriidés (Aves, Charadrii) در سایت Ramsar du Maroc: La Merja Zerga. گاو نر l’Institut Sci.-Fique 2006 ، 28 ، 35-47. [ Google Scholar ]
- مک کنزی، ال جی؛ Finkbeiner، MA; کرکمن، اچ. روش ها برای نقشه برداری توزیع علف های دریایی. در روش های تحقیق جهانی علف دریایی ; Elsevier Science BV: آمستردام، هلند، 2001; جلد 33، ص 101–121. [ Google Scholar ]
- آندراد، اف. Ferreira, MA روشی برای نظارت بر علفزارهای کم عمق دریایی ( Zostera spp.) با استفاده از عکاسی مایل زمینی در مقیاس بزرگ. آکوات. ربات. 2011 ، 95 ، 103-109. [ Google Scholar ] [ CrossRef ]
- Calleja، F. گالوان، سی. Silió-Calzada، A. Juanes، JA; Ondiviela، B. تجزیه و تحلیل بلند مدت Zostera noltei: یک رویکرد گذشته نگر برای درک پویایی علف های دریایی. مار. محیط زیست. Res. 2017 ، 130 ، 93-105. [ Google Scholar ] [ CrossRef ]
- کوهلوس، جی. استلزر، ک. مولر، جی. Smollich، S. نقشه برداری از علف دریایی (Zostera) با سنجش از دور در دریای Schleswig-Holstein Wadden. استوار. ساحل. Shelf Sci. 2020 , 238 , 106699. [ Google Scholar ] [ CrossRef ]
- تراگانوس، دی. Reinartz، P. تشخیص تغییر سالانه علفهای دریایی مدیترانه با استفاده از سری زمانی تصویر RapidEye. جلو. علوم گیاهی 2018 ، 9 ، 96. [ Google Scholar ] [ CrossRef ] [ نسخه سبز ]
- ژانگ، سی. استفاده از تکنیکهای همجوشی دادهها برای نقشهبرداری و پایش زیستگاه اعماق دریا در اکوسیستم صخرههای مرجانی. ISPRS J. Photogramm. Remote Sens. 2015 ، 104 ، 213-223. [ Google Scholar ] [ CrossRef ]
- تراگانوس، دی. Reinartz، P. نقشه برداری از علف های دریایی مدیترانه با تصاویر Sentinel-2. مارس آلودگی. گاو نر 2018 ، 134 ، 197-209. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- ژانگ، سی. سلچ، دی. زی، ز. رابرتز، سی. کوپر، اچ. چن، جی. نقشهبرداری زیستگاه اعماق دریا مبتنی بر شی در فلوریدا کیز از تصاویر فراطیفی. استوار. ساحل. Shelf Sci. 2013 ، 134 ، 88-97. [ Google Scholar ] [ CrossRef ]
- گام، م. بذری، ح. De Montaudouin, X. Impact de la présence d’herbiers à Zostera noltii sur l’infestation parasitaire des coques Cerastoderma edule dans la lagune de Merja Zerga (Maroc). گاو نر l’Institut Sci. 2009 ، 31 ، 13-20. [ Google Scholar ]
- گریگنون-دوبوآ، ام. Rezzonico، B. شیمی فنلی علف دریایی Zostera noltei Hornem. بخش 1: اولین شواهد از سه شیمیتیپ فلاونوئیدی زیرگونه در سه منطقه جغرافیایی متمایز. فیتوشیمی 2018 ، 146 ، 91-101. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- ناطیج، ل. خلیل، ک. لودیکی، م. Elkalay، K. اولین تلاش برای تقسیم مجدد علف های دریایی در سواحل مراکش. بین المللی J. Innov. علمی 2014 ، 10 ، 2351-8014. [ Google Scholar ]
- تهامی، ف. بذری، ح. بدوی، بی. بوارور، او. تالاب بنهوسا، آ. مرجا زرگا: مطالعه ساختار عملکردی و ارزیابی زیستی کیفیت اکولوژیکی جوامع کفزی. جی. ماتر. محیط زیست علمی 2017 ، 8 ، 4591-4599. [ Google Scholar ] [ CrossRef ]
- تهامی، ف. بذری، ح. بدوی، بی. Benhoussa، A. توزیع عمودی ماکروفون اعماق دریا در زیستگاه های جزر و مدی پراکنده شده توسط پرندگان ساحلی در تالاب مرجا زرگا. تالاس. بین المللی J. Mar. Sci. 2017 ، 34 ، 255-265. [ Google Scholar ] [ CrossRef ]
- پروئنسا، بی. فراپارت، اف. لوباک، بی. ماریو، وی. ایگورا، بی. بومبرون، ال. میشالت، آر. سوتولیچیو، الف. پتانسیل تصاویر با وضوح بالا برای نظارت بر تکامل مرداب نمکی پس از تهاجم اسپارتینا. Remote Sens. 2019 , 11 , 968. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- واله، ام. پالا، وی. لافون، وی. Dehouck، A. Garmendia, JM; بورجا، Á. Chust، G. نقشه برداری زیستگاه های مصب رودخانه با استفاده از تصاویر فراطیفی هوابرد، با تمرکز ویژه بر علفزارهای علف دریایی. استوار. ساحل. Shelf Sci. 2015 ، 164 ، 433-442. [ Google Scholar ] [ CrossRef ]
- تولدال، اچ ام. Wikström، SA طبقه بندی پوشش گیاهی و بسترهای آبی با لیدار هوابرد. سنسور از راه دور محیط. 2012 ، 121 ، 347-357. [ Google Scholar ] [ CrossRef ]
- پان، ز. فرناندز-دیاز، جی سی. گلنی، CL; استارک، ام. علف دریایی آب کم عمق مشاهده شده توسط LiDAR شکل موج کامل با وضوح بالا. در مجموعه مقالات سمپوزیوم IEEE Geoscience and Remote Sensing 2014، شهر کبک، QC، کانادا، 13 ژوئن 2017. [ Google Scholar ]
- زاوالاس، ر. ایرودیاکونو، دی. رایان، دی. راتری، ا. Monk, J. طبقهبندی زیستگاه جوامع درشت جلبکی دریایی معتدل با استفاده از LiDAR Bathymetric. Remote Sens. 2014 , 6 , 2154–2175. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- ایشی گورو، اس. یامادا، ک. یاماکیتا، تی. یامانو، اچ. اوگوما، اچ. Matsunaga، T. طبقهبندی بسترهای علف دریایی با جفت کردن دادههای عمق سنجی LiDAR هوابرد و عکسهای هوایی دیجیتال. در CO 2 ، دما، و درختان . Springer: سنگاپور، 2016; صص 59-70. [ Google Scholar ]
- پریش، م. Dijkstra، JA; اونیل-دان، JPM؛ مک کنا، ال. Pe’eri, S. نقشهبرداری زیستگاه اعماق دریا پس از سندی با استفاده از فناوری جدید lidar topobathymetric و طبقهبندی تصویر مبتنی بر شی. جی. ساحل. Res. 2016 ، 76 ، 200-208. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- وبستر، تی. مک گویگان، ک. کراول، ن. کالینز، کی. Macdonald, C. بهینهسازی جمعآوری دادهها و اصلاح تکنیکهای پس پردازش برای اولین بررسی توپوگرافی آبهای کم عمق Lidar. جی. ساحل. Res. 2016 ، 76 ، 31-43. [ Google Scholar ] [ CrossRef ]
- وبستر، تی. نتایج از 3 فصل بررسی در دریای کانادا با استفاده از حسگر لیدار آب کم عمق Leica Chiroptera II. In Proceedings of the OCENS 2017، آبردین، بریتانیا، 19 تا 22 ژوئن 2017؛ صص 1-4. [ Google Scholar ] [ CrossRef ]
- کولینگز، اس. کمبل، NA; Keesing، JK کمی سازی قدرت تبعیض آمیز فناوری های سنجش از راه دور برای نقشه برداری زیستگاه اعماق دریا. بین المللی J. Remote Sens. 2018 , 40 , 2717–2738. [ Google Scholar ] [ CrossRef ]
- Dierssen, HM; Zimmerman، RC; چرم، RA; داونز، تلویزیون؛ دیویس، CO سنجش از دور رنگی اقیانوس از علفهای دریایی و آبسنجی در بانکهای باهاما با تصاویر هوایی با وضوح بالا. لیمنول اقیانوسگر. 2003 ، 48 ، 444-455. [ Google Scholar ] [ CrossRef ]
- گارسیا، RA; هدلی، جی دی. قلع، HC; Fearns، PRCS روشی برای تجزیه و تحلیل پتانسیل سنجش از دور نوری برای نقشه برداری زیستگاه های اعماق دریا. Remote Sens. 2015 ، 7 ، 13157–13189. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- پان، ز. گلنی، سی. دیاز، JCF; استارک، ام. مقایسه حمامسنجی و نقشهبرداری علفدریایی با تصاویر فراطیفی و لیدار عمقسنجی هوابرد در یک محیط مصب کم عمق. بین المللی J. Remote Sens. 2016 , 37 , 516–536. [ Google Scholar ] [ CrossRef ]
- کاستیو لوپز، ای. دومینگوئز، جی. پردا، ر. دی لوئیس، جی.ام. پرز، آر. پینا، اف. اهمیت تصحیح جوی برای سنجش از دور ابرطیفی هوابرد آبهای کم عمق: کاربرد در تخمین عمق. اتمس. Meas. فنی 2017 ، 10 ، 3919–3929. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- فردریکسن، م. کراوس-جنسن، دی. هولمر، ام. Laursen، JS تغییرات مکانی و زمانی در مناظر علف مارماهی (Zostera marina): تأثیر محیط فیزیکی. آکوات. ربات. 2004 ، 78 ، 147-165. [ Google Scholar ] [ CrossRef ]
- Lathrop، RG; مونتسانو، پی. Haag, S. یک رویکرد تقسیم بندی چند مقیاسی برای نقشه برداری زیستگاه های علف دریایی با استفاده از تصاویر دوربین دیجیتال هوابرد. فتوگرام مهندس Remote Sens. 2006 ، 72 ، 665-675. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- فلچر، RS; پولیچ، دبلیو. هاردگر، بی. یک رویکرد نیمه خودکار برای نظارت بر تغییرات چشم انداز در بسترهای علف دریایی تگزاس از عکاسی هوایی. جی. ساحل. Res. 2009 ، 252 ، 500-506. [ Google Scholar ] [ CrossRef ]
- جوان، DR. کلینتون، پی جی؛ Specht، DT نقشه برداری از علف مارماهی جزر و مدی ( Zostera marina L.) در سه مصب ساحلی اقیانوس آرام شمال غربی ایالات متحده آمریکا با استفاده از تصویربرداری هوایی نزدیک به مادون قرمز رنگ کاذب. بین المللی J. Remote Sens. 2010 ، 31 ، 1699-1715. [ Google Scholar ] [ CrossRef ]
- Uhrin، AV; Townsend، PA با استفاده از اختلاط طیفی خطی عکس های هوایی، نقشه برداری از علف های دریایی را بهبود بخشید. استوار. ساحل. Shelf Sci. 2016 ، 171 ، 11-22. [ Google Scholar ] [ CrossRef ]
- گالستروم، ام. لوندن، بی. بودین، م. کانگوه، جی. اوهمان، ام سی; Mtolera، MS; Björk, M. ارزیابی تغییرات در پوشش گیاهی غوطه ور تحت سلطه علف های دریایی خلیج چواکا استوایی (زانزیبار) با استفاده از سنجش از دور ماهواره ای. استوار. ساحل. Shelf Sci. 2006 ، 67 ، 399-408. [ Google Scholar ] [ CrossRef ]

شکل 1. ایستگاه های نمونه برداری Z. noltei در تالاب مرجا زرگا: S1, S2, S3, S4, S5, S6, S7 (منبع: Image SPOT 7).
شکل 2. نمودار جریان پردازش داده ها برای نقشه برداری از علف مارماهی کوتوله Z. noltei و جلبک در تالاب Merja Zerga.
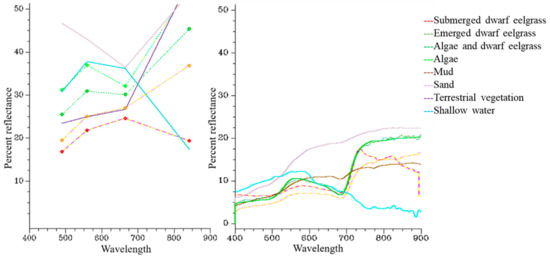
شکل 3. نشانه های طیفی برای هشت نوع زیستگاه مختلف در تالاب مرجا زرگا: Z. noltei ظهور ، Z. noltei غوطه ور ، جلبک، بستر مخلوط Z. noltei و جلبک، ماسه، گل و آب کم عمق. سمت چپ: اعداد طیف متوسط (n = 5) به چهار باند تصویر ماهوارهای SPOT 7 تنزل یافته است: 1 (آبی؛ 450-520 نانومتر)، 2 (سبز؛ 530-590 نانومتر)، 3 (قرمز؛ 625-695 نانومتر)، و 4 (NIR؛ 760-890 نانومتر). راست: طیف متوسطی که در وضوح طیفی طیفسنج JAZA0478 به دست میآید.

شکل 4. کوادرات های Z. noltei در ایستگاه همگن ( a ) S3 و در ایستگاه های مختلط ( b ) S5 (متراکم) و ( c ) S7 (پراکنده، غوطه ور)، عکاسی بستر مربوطه آنها ( d – f ).
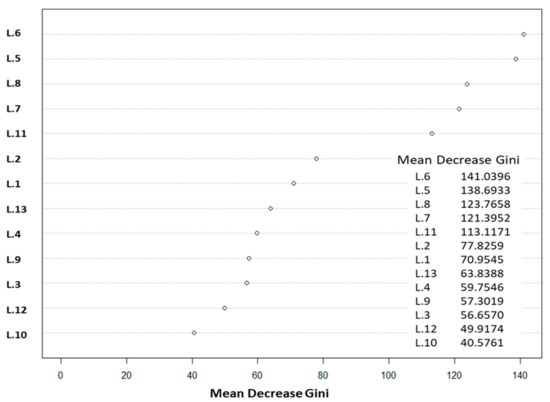
شکل 5. نتایج تصادفی جنگل: میانگین کاهش مقادیر جینی برای 13 باند (L.1: باند 1؛ L.2: باند 2؛ L.3: باند 3؛ L.4: باند 4؛ L.5: باند 1. L.6: NDVI؛ L.7: CVI؛ L.8: GVI؛ L.9: ICS؛ L.10: NLI؛ L.11: EVI؛ L.12: IB؛ L.13: SAVI).
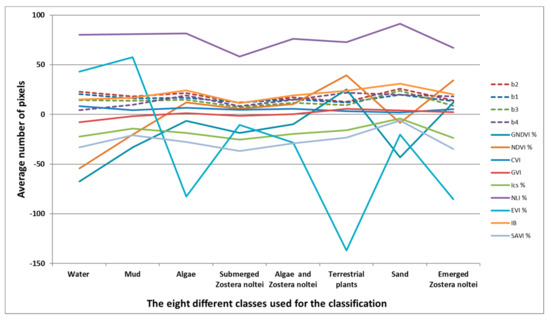
شکل 6. میانگین 1371 پیکسل در هر کلاس برای هر یک از 4 باند و 9 شاخص تصویر SPOT 7.
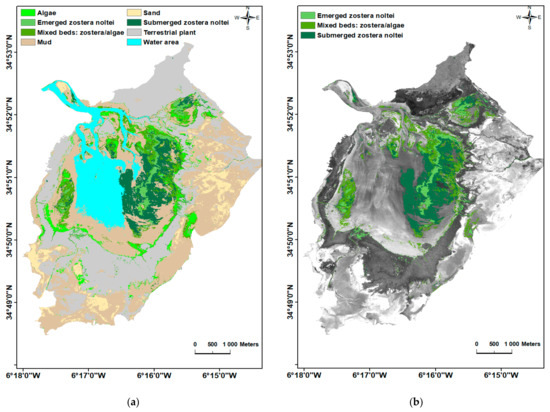
شکل 7. ( الف ) نتایج طبقهبندی تصادفی جنگل و ( ب ) توزیع علف مارماهی کوتوله Z. noltei و جلبکهای بزرگ در تالاب Moulay Bousselham.
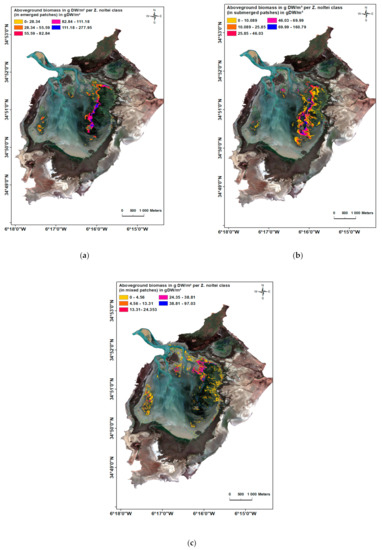
شکل 8. زیست توده روی زمین بر حسب گرم DW/m2 به ازای کلاس Z. noltei ( a – c ) در تالاب Merja Zerga.

شکل 9. گونههای جلبک بزرگ مرتبط با Z. noltei ( a ) و ماکروجلبکهای سبز رشتهای ( b ) در بسترهای مخلوط مشاهدهشده در ناحیه جزر و مدی تالاب Merja Zerga.


بدون دیدگاه