

نقش حداکثر آنتروپی و علم شهروندی در مطالعه تناسب زیستگاه فاخته ژاکوبین در سناریوهای مختلف تغییر اقلیم
چکیده
پیشرفتهای اخیر در روشهای مدلسازی و نقشهبرداری فضایی، افقهای جدیدی را برای نظارت بر مهاجرت گونههای پرندگان، که به دلیل تغییرات آب و هوایی تغییر کردهاند، باز کرده است. ظهور علم شهروندی همچنین به جمع آوری داده های مکانی و زمانی با ویژگی های مرتبط کمک کرده است. دادههای تنوع زیستی از رصدخانههای شهروندان را میتوان در الگوریتمهای یادگیری ماشینی برای پیشبینی شرایط محیطی مناسب برای بقای گونهها و رفتارهای مهاجرت آتی آنها به کار برد. در این مطالعه، متغیرهای محیطی مختلف موثر در مهاجرت پرندگان مورد تجزیه و تحلیل قرار گرفت و مناسب بودن زیستگاه آنها برای درک آتی از پاسخ آنها در سناریوهای مختلف تغییر اقلیم ارزیابی شد. فاخته ژاکوبین ( Clamator jacobinus) به عنوان گونه موضوع انتخاب شد، زیرا ورود آنها به هند به طور سنتی به عنوان نشانه ای برای شروع فصل باران های موسمی هند در نظر گرفته می شود.

برای پیشبینی مناسب بودن در سناریوهای فعلی و آینده، مدلسازی حداکثر آنتروپی (Maxent) با متغیرهای محیطی و وقوع گونههای مشاهدهشده در هند و آفریقا انجام شد. برای مدل سازی، آزمون همبستگی بر روی متغیرهای محیطی (زیست اقلیم، بارش، حداقل دما، حداکثر دما، بارش، باد و ارتفاع) انجام شد. نتایج نشان داد که متغیرهای مرتبط با بارش نقش مهمی در تناسب داشتند و از طریق نقشههای مناسب زیستگاهی طبقهبندیشده، مشاهده شد که مناطق مناسب هند و آفریقا ممکن است در سناریوهای اقلیمی آینده (SSPs 2.6، 4.5، 7.0 و 8.5) کاهش یابد. 2030 و 2050. علاوه بر این،2 ) مشاهده تغییرات ظریف در اکوسیستم. چنین مطالعات تغییرات آب و هوایی می تواند از تحقیقات تنوع زیستی حمایت کرده و اقتصاد کشاورزی را بهبود بخشد.
کلید واژه ها:
دانش شهروندی ; یادگیری ماشینی ؛ باران های موسمی هند ؛ فاخته ژاکوبن ; مکسنت ؛ مدل توزیع گونه ; مناسب بودن زیستگاه ; گسترش دامنه ; WorldClim ; CMIP
1. مقدمه
تغییر تصاعدی در اقلیم به طور مستقیم بر توزیع فضایی گونهها و جوامع در اکوسیستمها تأثیر گذاشته است، که یک نیاز ضروری برای درک عملکردها و فرآیندهای اکوسیستم است. به این ترتیب، حرکت گونهها در پاسخ به تغییرات اقلیمی میتواند از مدلهای توزیع گونهها (SDMs) پیشبینی شود، که روشی تجربی برای ارزیابی اثرات اقلیمی برای تغییرات زیستگاه گونهها ارائه میکند (به عنوان مثال، مرجع [ 1 ]). زیستگاه به عنوان مکان خاصی تعریف می شود که در آن گونه ها با ویژگی ها، رفتار، تعاملات و الگوهای جمعیتی معین زندگی و تولید مثل می کنند [ 2 ]. زیستگاه مطلوبی که برای بقای یک گونه مهم است، مناسب بودن زیستگاه نامیده می شود [ 3] و در تحقیقات اکولوژیکی از طریق مدل سازی مناسب زیستگاه اهمیت دارد که می تواند در برنامه های حفاظتی و حفاظتی کمک کند. چندین مطالعه مناسب بودن زیستگاه گونه ها را با استفاده از حداکثر آنتروپی (Maxent) [ 4 ] برای ارزیابی محدوده گونه ها با استفاده از داده های موقعیت جغرافیایی بررسی کرده اند. به عنوان مثال، مراجع [ 5 ، 6 ، 7 ، 8 ، 9 ، 10 ، 11 ، 12 ]. مدل Maxent در ادبیات برای مدلسازی توزیعهای فضایی گونهها محبوبیت پیدا کرده است، و مطالعات مرتبط بیش از 5000 استناد در مجموعه Web of Science Core دریافت کردهاند که بیشتر توسط مدلسازان توزیع (حدود 60 درصد) استفاده میشود [ 13 ].]. در دهه های اخیر، مطالعات بی شماری در مورد مناسب بودن زیستگاه گونه ها با استفاده از روش حداکثر آنتروپی برای ارزیابی محدوده گونه ها با استفاده از داده های فقط حضوری انجام شده است [ 14 ، 15 ، 16 ، 17 ، 18 ، 19 ]. چنین مطالعاتی به استخراج دستورالعملهای مفید در مورد پارامترهای مدل، مانند حداقل حجم نمونه و نیاز به نمونه داده، انتخاب نمونههای تصادفی از مجموعه دادههای حجیم و تعیین فرآیند نمونهگیری فرعی برای پیشبینی محدوده در هر گونه و هر حجم نمونه کمک میکند [ 14 ، 15 ، 16 ، 18 ، 19 ].
اگرچه چنین رویکردهای مدلسازی مبتنی بر دادهها میتواند به شناسایی عوامل اصلی توسعه دامنه گونهها کمک کند [ 20 ]]، وقوع گونهها، که سهم قابل توجهی در مدل دارند، ترجیحاً باید در زمینههای مکانی (محدوده گونهها) و زمانی (زمان مشاهده) ثبت شوند. با این حال، ذاتاً یک کار مشکل ساز و پرهزینه برای ثبت مشاهدات جغرافیایی متنوع در زمان واقعی است، زیرا چنین فعالیت هایی نیاز به نظارت مداوم بر حرکات گونه ها دارند، بنابراین گونه ها با دستگاه های ردیابی برچسب گذاری می شوند بدون اینکه آسیبی به آنها وارد شود. بنابراین، این مشاهدات زمان واقعی یا نزدیک به زمان واقعی از طریق یک رویکرد داوطلبانه برای جمعآوری دادهها میتواند به کمی کردن تناسب گونهها در مقیاس فضایی بزرگ و اطلاعرسانی در مورد تغییرات در الگوهای اقلیمی کمک کند. خواص گونه را می توان از گیاهان، ادبیات،21 ]. با این حال، چالش اصلی همچنان عدم قطعیت فضایی است که ممکن است از برچسبگذاری جغرافیایی نادرست یا اطلاعات مبدأ اشتباه ناشی شود [ 21 ، 22 ]. تا همین اواخر، دادههای گونهها بهعنوان توصیف متنی در قالب نامها و مکانها [ 22 ] جمعآوری و ثبت میشد و دیجیتالی کردن اطلاعات متنی نیز باعث خطاهای اساسی میشود و عدم قطعیت فضایی در حد چند کیلومتر را به همراه میآورد [ 23 ]. تکنیکهای مختلفی برای تخمین و مستندسازی عدم قطعیت مکان در میان سوابق وقوع گونهها به منظور حذف خطاهای بالا قبل از مدلسازی مناسب توسعه داده شده است [ 22 ، 24 ].
داوطلبان آموزش دیده و آموزش ندیده در فرآیندهای جمع آوری داده ها به عنوان یک رویکرد علم شهروندی کمک کرده اند، که ممکن است داده های قوی و دقیق با ویژگی های کیفی و کمی ارائه دهد. دادههای جمعآوریشده با استفاده از رویکردهای علم شهروندی در سالهای اخیر برای کاهش شکافها در کمیت و کیفیت دادهها برای مدلهای تخصصی اکولوژیکی اعمال شدهاند، که همچنین تقریب معیار مورد علاقه را بهبود بخشید [ 25 ]. مدلسازی توزیع گونهها برای گونههای خاص به تعداد کافی از رخدادها نیاز دارد که در سراسر موجود آن توزیع شده است [ 26 ، 27 ، 28 ]. علم شهروندی مفهومی گسترده است که به اشکال مختلف قابل درک است [ 29]، از پروتکل های بسیار سیستماتیک گرفته تا بررسی های فرصت طلبانه بدون طرح های نمونه [ 29 ، 30 ].
در این مقاله، یک الگوریتم حداکثر آنتروپی مبتنی بر ML (Maxent) برای رخدادهای فاخته ژاکوبین (فاخته یا فاخته کاکل دار) با متغیرهای محیطی، مانند ارزیابی مناسب بودن زیستگاه بالقوه در آفریقا و هند، اعمال شد. برای روش مدلسازی، وقوع پرندگان ابتدا به سه دوره زمانی تقسیم شد – ژوئن تا سپتامبر، که به موسمی جنوب غربی هند اشاره دارد، اکتبر تا دسامبر، که به موسمی شمال غرب هند و فصل مرطوب آفریقا اشاره دارد، و ژانویه تا مه، برای پیشبینی. محل مناسب بودن فاخته ژاکوبن است، زیرا اکثر مناطق هند دارای فصل زمستان و تابستان هستند. این رویکرد همچنین برای پیشبینی تغییر الگوهای موسمی با مدلسازی چگونگی تغییر زیستگاههای مطلوب این پرنده تحت سناریوهای مختلف تغییر آب و هوا مورد استفاده قرار گرفت. برای این مدل سازی پیش بینی آینده، نتایج مدلسازی بهدستآمده از تناسب فعلی با متغیرهای محیطی موجود و وقوع گونهها با مدلهای آینده برای مشاهده تغییرات آب و هوایی احتمالی در سالهای 2030 (میانگین 2021-2040) و 2050 (میانگین 2041-2060) پیشبینی شد. علاوه بر این، مناطق مناسب و نامناسب بودن سایت ها نیز برای تجزیه و تحلیل افزایش یا کاهش در سیستم اکولوژیکی در پاسخ به تغییرات در الگوهای موسمی محاسبه شد. اولین مطالعه تغییرات آب و هوایی موسمی هند از نظر مهاجرت فاخته های ژاکوبین توسط سینگ و ساران انجام شد. مناطق مناسب و نامناسب بودن سایت ها نیز برای تجزیه و تحلیل افزایش یا کاهش در سیستم اکولوژیکی در پاسخ به تغییرات در الگوهای موسمی محاسبه شد. اولین مطالعه تغییرات آب و هوایی موسمی هند از نظر مهاجرت فاخته های ژاکوبین توسط سینگ و ساران انجام شد. مناطق مناسب و نامناسب بودن سایت ها نیز برای تجزیه و تحلیل افزایش یا کاهش در سیستم اکولوژیکی در پاسخ به تغییرات در الگوهای موسمی محاسبه شد. اولین مطالعه تغییرات آب و هوایی موسمی هند از نظر مهاجرت فاخته های ژاکوبین توسط سینگ و ساران انجام شد.31] که در آن رخدادهای جغرافیایی فاخته ژاکوبین با 19 متغیر زیست اقلیم فعلی با استفاده از مدل Maxent مبتنی بر ML در نرم افزار R مدل سازی شد. سپس این مدل آموزشدیده با متغیرهای زیستاقلیمی آینده تحت سناریوی RCP8.5 پروژه مقایسهای مدل جفت شده (CMIP5) برای ارزیابی تغییرات پیشبینیشده در مناسب بودن زیستگاههای فاختههای Pied تا سال 2050 پیشبینی شد. به طور خاص، متغیرهای زیستاقلیمی فعلی و آینده در وضوح 2.5 ثانیه قوسی (~4.5 کیلومتر در خط استوا) طول و عرض جغرافیایی استفاده می شود. مطالعه فوق نشان داد که متغیرهای اصلی محیطی که بر مناسب بودن فاخته ژاکوبین تأثیر میگذارند همدما (8/16 درصد)، میانگین دمای گرمترین فصل (7/15 درصد)، بارش سالانه، بارش گرمترین سهماهه (13) میباشد. 6٪ و بارش مرطوب ترین ماه (11.3٪) در طول فصل باران های موسمی تابستان هند، یعنی ژوئن تا اکتبر. بر اساس پیشبینیهای کنونی مناسببودن، ایالتهای جنوب هند – آندرا پرادش، گوا، کارناتاکا، کرالا، ماهاراشترا و تامیل نادو – و شمال هند – اوتاراکند و هیماچال پرادش – تناسب زیستگاهی بالا و همچنین مناسب زیستگاه متوسط و غرب را نشان دادند. ایالات (به عنوان مثال، گجرات) مناسب بودن متوسط را نشان داد. جنوب آفریقا برای این پرنده نامناسب تشخیص داده شد، زیرا در ماه های ژوئن تا اکتبر، آب و هوای خشک و گرم در آنجا تجربه می شود که زیستگاه مطلوبی نیست. با این حال، با توجه به پیش بینی مناسب بودن آینده این پرنده، انقباض محدوده فاخته ژاکوبین می تواند در تمام نقاط هند به جز بخش های جنوبی تامیل نادو به دلیل افزایش انتشار گازهای گلخانه ای و کاهش بارندگی در گرم ترین سه ماهه اتفاق بیفتد. علاوه بر این، چندک (5٪ و 95٪) از متغیرهای محیطی مربوطه برای مشاهده تغییرات آب و هوای کنونی و 2050 با توجه به فصول موسمی هند محاسبه شد.
2. علم شهروندی به عنوان روش تحقیق تنوع زیستی
عمل درگیر کردن داوطلبان در کارهای علمی در چند دهه گذشته با فرصتهای مبرمتر ارائهشده برای شرکتکنندگان برای ارائه رویکردهای پیشرفته و ایجاد بینش معنادار در مورد دادههای جمعآوریشدهشان، افزایش یافته است. فعالیت استفاده موثر از جمع سپاری، همراه با اینترنت و برنامه های کاربردی تلفن همراه، در مناطق جغرافیایی بزرگ به عنوان علم شهروندی شناخته می شود. علم شهروندی «فرآیندی است که در آن شهروندان، سازمانهای دولتی، صنعت، دانشگاه، گروههای اجتماعی و مؤسسات محلی برای نظارت، پیگیری و پاسخگویی به مسائل مربوط به نگرانی مشترک جامعه همکاری میکنند» [ 32 ] و «جایی که شهروندان و ذینفعان در مدیریت منابع» [ 33 ، 34]. علم شهروندی شامل افراد حرفهای و غیرحرفهای است که هم در تفکر علمی و هم در جمعآوری دادهها [ 33 ، 35 ] با پشتیبانی از پیشرفتهای فناوری، مانند تلفنهای هوشمند با خدمات مکانیابی، دوربین، شتابسنج و غیره شرکت میکنند [ 36 ]. با این حال، علم شهروندی بر اساس ماهیت مشارکت و سودمندی آن در حوزههای مختلف ممکن است تعاریف و معانی مفهومی متفاوتی داشته باشد.
با توجه به ماهیت تعامل، کهکشان علم شهروندی به چهار سطح طبقهبندی میشود: جمعسپاری، هوش توزیعشده، علم مشارکتی و افراطی [ 37 ]. جمع سپاری ابتدایی ترین سطحی است که عموم مردم می توانند با پردازش و تجزیه و تحلیل داده های جمع آوری شده به علم کمک کنند. سطح بعدی هوش توزیع شده است که در آن شهروندان قبل از مشارکت در فعالیتهای جمعآوری و تفسیر دادهها، مهارتهای جدیدی را یاد میگیرند. سطح سوم علم مشارکتی است که در آن شهروندان با گروه های تحقیقاتی برای تعریف مشکلات و جمع آوری داده ها درگیر می شوند. آخرین سطح افراطی است، جایی که شهروندان به کنترل کامل برای تعریف مشکلات، جمع آوری داده ها و انجام تجزیه و تحلیل بر روی آن مجهز هستند.
ویگینز و کرونستون [ 38 ] با توجه به کاربرد آن در پروژه های مختلف با اهداف مختلف، پروژه های علمی شهروندی را به پنج نوع منحصر به فرد و جامع متقابل طبقه بندی کردند: اقدام، آموزش، حفاظت، تحقیق و پروژه های مجازی. پروژه های اقدام مختلف با همکاری مشترک شهروندان و دانشمندان/محققان به مسائل محلی می پردازند – برای مثال، منابع [ 39 ، 40 ، 41 ، 42 ، 43 ، 44 ، 45 ، 46 ، 47 ، 48 ، 49 ] و پروژه های آموزشی کمک می کنند. در ارتقای دانش شهروندان به عنوان بخشی از برنامه درسی [50 , 51 , 52 , 53 , 54 , 55 ]. پروژه های حفاظتی بر مدیریت منابع طبیعی متمرکز هستند – برای مثال، مرجع [ 19 ] – پروژه های تحقیقاتی بر مطالعه مشاهدات شهروندان همراه با پارامترهای مختلف برای پاسخ به سوالات علمی [ 56 ، 57 ] تاکید دارند و پروژه های مجازی شامل فعالیت های علمی شهروندی از راه دور است [ 58 ]. , 59 , 60 , 61 ].
طرحهای طبقهبندی فوق را میتوان با مثال Project PigeonWatch نشان داد، که یکی از پروژههای علمی شهروندی در آزمایشگاه پرندهشناسی کورنل (CLO) و انجمن ملی Audubon است که بسیاری از داوطلبان از همه سنین و حرفهها را در سراسر جهان برای جمعآوری درگیر میکند. داده های عملی برای مطالعه و تجزیه و تحلیل تغییرات رنگ کبوتر. بر اساس چک لیست فوق، این پروژه را می توان به عنوان یک پروژه “تحقیق” توصیف کرد و رویکرد مورد استفاده “جمع سپاری” است.
در میان پروژههای علمی شهروندی مختلف، 72 درصد از پروژهها به رشته زیستشناسی [ 62 ] مربوط میشود و به دلیل چنین تسلطی در این حوزه، مطالعه و تحقیق در مورد تنوع و پراکنش گونهها [ 63 ] نیاز سریع به تنوع زیستی را پیش میبرد. نظارت، برنامه ریزی حفاظت و تحقیقات زیست محیطی. بسیاری از برنامههای علوم شهروندی در طول سالها یا حتی دههها محقق شدهاند و هنوز هم برای مطالعه الگوهای طبیعت در مقیاس فضایی بزرگ با جمعآوری دادهها در مورد مکانها و زیستگاههای مختلف گونهها انجام میشوند. نحوه جمع آوری چنین اطلاعاتی در مورد مکان گونه ها، زیستگاه ها و سایر اطلاعات مرتبط [ 63 ، 64 ، 65] با جذب عموم مردم در فعالیت های علمی در حال حاضر بهترین روش تلقی می شود. لزوماً درست نیست که خروجی علمی همیشه از راهبردها و استنباطهای قوی از انتشارات علمی بسیار شناخته شده و مورد بازبینی بهره میبرد. در عوض، جمع آوری اطلاعات از طریق مشارکت عمومی می تواند منبع بهتری از اطلاعات علمی برای پاسخ به سؤالات خاص باشد [ 66 ، 67 ]. ممکن است اعتبار بیشتری به آزمایشگاه پرنده شناسی دانشگاه کرنل داده شود که پایه و اساس مشارکت داوطلبانه در مشاهدات، نظارت و تحقیقات تنوع زیستی را ایجاد کرد [ 52 ]]. با این حال، بسیاری از سازمانها و گروههای تحقیقاتی دیگر نیز وجود دارند که برنامههای علوم شهروندی مختلفی را برای جمعآوری دادههای جغرافیایی خوب و متراکم با نمونهگیری مکانی دقیق طراحی کردهاند، مانند نقشهبرداری گونهها از طریق پورتال شبکه اطلاعات منابع زیستی هند (IBIN) [ 68 ]، bioblitz. [ 69 ]، بررسی پلی مورفیسم پوسته [ 70 ]، بررسی کیفیت آب [ 71 ] و بررسی پرندگان پرورشی [ 72 ]. چنین مجموعه دادههای متنوعی، تجمیع دادههای مشاهده از منابع مختلف را برای تحقیقات مرسوم مجبور میکند، اما نگرانیهای عمده حتی پس از تجمیع دادهها، کیفیت داده [ 73 ] و تکنیکهایی برای ترکیب مجموعههای داده متنوع در طرحوارههای مختلف است [ 74 ]]. بنابراین، برنامهریزی مناسبی برای مدیریت یکپارچهسازی مجموعه دادههای حجیم در طرحی یکنواخت با زیرساخت بررسی کیفیت داده برای مدیریت سوگیریهای مشاهدهای، «غیابهای کاذب» که منجر به مشاهدههای ناکافی [ 75 ] و توزیع نابرابر داده [ 76 ] میشود، مورد نیاز است. این چالش ها توسط یک تلاش هماهنگ جهانی [ 77 ] که در سال 2004 آغاز شد و اکنون به بزرگترین دروازه واحد به مجموعه داده های مبتنی بر مشاهده، معروف به تسهیلات جهانی اطلاعات تنوع زیستی (GBIF) منجر شده است، مورد توجه قرار گرفت.
GBIF یک سازمان بیندولتی است که «یک شبکه قابل دسترسی به اینترنت و قابلیت تعامل از پایگاههای اطلاعاتی تنوع زیستی و ابزارهای فناوری اطلاعات» [ 78 ] را بهعنوان «منبع سنگ بنا» [ 79 ]، با «مأموریت در دسترس قرار دادن دادههای تنوع زیستی جهان بهصورت آزاد و جهانی» فراهم میکند. از طریق اینترنت» [ 79 ]. در حال حاضر، پورتال GBIF دسترسی آزاد به بیش از 160 میلیون رویداد تنوع زیستی و داده های گونه از 1641 موسسه و داده های نقشه برداری داوطلبانه در سراسر جهان را فراهم می کند. بنابراین، GBIF به یک مخزن معتبر تبدیل شده است که در آن سازمان ها/موسسات مختلف داده های خود را با کیفیت و به مقدار زیاد به اشتراک می گذارند، که برای اهداف مدل سازی و تصمیم گیری ضروری است. ادواردز و همکاران [ 77] اعتبار فضایی سومین خانواده بزرگ گیاهان گلدار، لگومینوزا را با استفاده از گونهها و دادههای توزیع آن از پورتال GBIF برای ارزیابی کیفیت و پوشش رخدادهای جغرافیایی آن انجام داد. نظرات مشابهی توسط گراهام و همکاران مشاهده شده است. [ 21 ] و سوارز و تسوتسوی [ 80 ] برای استفاده های اضافی از داده های نمونه موزه، که سیاست تنوع زیستی و فرآیند تصمیم گیری را تسهیل می کرد [ 80 ]. در میان مزایای مختلف دیگر، داده های GBIF را می توان برای ارزیابی تنوع زیستی [ 81 ]، تجدید نظرهای طبقه بندی [ 82 ]، گردآوری لیست های قرمز گونه های در معرض تهدید [ 83 ] و مدل سازی مناسب زیستگاه استفاده کرد [ 31 ، 84 ،85 ، 86 ، 87 ]. مورد دوم یکی از نمونه های برجسته مطالعات تغییرات آب و هوایی است که در آن مشاهدات مبتنی بر علم شهروندان از GBIF به طور فزاینده ای مورد استفاده قرار می گیرد [ 88 ، 89 ، 90 ، 91 ، 92 ، 93 ، 94 ، 95 ]. در این مقاله، سناریوهای مختلف تغییر اقلیم همراه با رخدادهای مشاهده شده توسط GBIF از پرنده مطلوب موسمی، Clamator jacobinus ، با استفاده از رویکرد Maxent برای مطالعه مناسب بودن زیستگاه معاصر و آینده این پرنده مدلسازی شدهاند تا تغییرات در فصل بارانهای موسمی هند بتواند مورد بررسی قرار گیرد.
3. مواد و روشها
3.1. گونه فاخته ژاکوبن (پایدار).
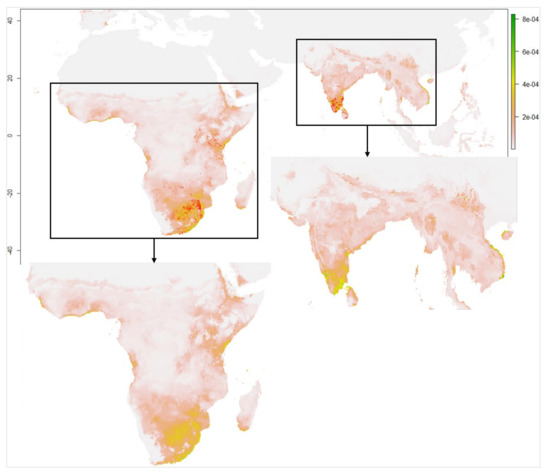
طبق باور هندی ها، ورود این پرنده نیمه مهاجر، فاخته ژاکوبین ( Clamator jacobinus ) ( شکل 1 )، همچنین به عنوان “چاتاک” در هند شناخته می شود، منادی شروع موسمی هند است [ 96 ]. همانطور که در شکل 2 نشان داده شده است، در طول تابستان، پرنده برای تولید مثل از آفریقا به هند پرواز می کند و از دریای عرب و اقیانوس هند عبور می کند. فاخته ژاکوبن متعلق به راسته فاختههای پرندگان کوچک زمینی با پرهای نرم سیاه و سفید و بالهایی بلند با تاجهای پفدار روی سر است که با قطرات باران تشنگی خود را برطرف میکند. این گونه به عنوان یک انگل مولد نیز شناخته می شود، یعنی به جای اینکه لانه خود را بسازد، تخم های خود را در لانه پرندگان دیگر، به ویژه خرچنگ جنگل ( Turdoides striata ) می گذارد.). این پرنده با طبیعت درختی بیشتر بر روی درختان بلند می نشیند اما اغلب در بوته های کم ارتفاع و گاهی اوقات روی زمین به دنبال غذا می گردد. مناطق جنگلی، جنگل ها و بوته ها را در مناطق نیمه خشک ترجیح می دهد. همانطور که به طور گسترده شناخته شده است، فاخته ژاکوبین از دو طریق مناسب بودن خود را در هند حفظ می کند. جمعیتی وجود دارد که به عنوان ساکن در تمام طول سال در بخش جنوبی کشور دیده می شود. فاخته ژاکوبین همچنین در بخشهای مرکزی و شمالی هند همراه با بادهای موسمی تابستانی از قبل از بارانهای موسمی تا اوایل زمستان، یعنی مه تا اوت، دیده میشود. دلیل انتخاب فاخته ژاکوبن این بود که زمان رسیدن این پرنده مستقیماً با موسمی مرتبط است زیرا فقط قطرات آب باران را مینوشد و از هیچ منبع آبی دیگری مانند آب باران، رودخانهها و غیره استفاده نمیکند.
3.2. داده های توزیع گونه ها و پیش پردازش
داده های توزیع از مخزن GBIF به دست آمده است که سوابق جغرافیایی این پرنده را از نظرسنجی ها، موزه ها، مشاهدات انسانی و سایر منابع داده جمع آوری می کند. سپس، موارد ثبت شده از طریق “مشاهده انسانی” برای این مطالعه انتخاب شدند، زیرا این تحقیق بر نشان دادن استفاده از داده های علوم شهروندی برای مدل سازی مناسب زیستگاه متمرکز بود [ 97 ]]. مؤسسات/سازمانها دادههای این پرنده را در دسته «مشاهده انسانی» از طریق برنامههای مختلف علمی شهروندی ارائه کردند. این مؤسسات/سازمانها عبارتند از: آزمایشگاه پرندهشناسی کرنل، مؤسسه پرندهشناسی آفریقای فیتز پاتریک، مؤسسه ملی تنوع زیستی آفریقای جنوبی، iNaturalist.org، Observation.org، بنیاد Xeno-canto برای صداهای طبیعت، naturgucker.de، سرویس حیات وحش کنیا، تنوع زیستی پورتال هند. و روچا کنیا. با این حال، مجموعه داده حاوی مقادیر مکرر طول و عرض جغرافیایی و همچنین مقادیر صفر بود (NA: در دسترس نیست). با استفاده از الگوریتم پاکسازی داده ها در R، رکوردهای دارای مقادیر NA و مکان های تکراری حذف شدند. از آنجایی که فاخته های ژاکوبین به دلیل ارتباط نزدیکشان با شروع فصل باران های موسمی در هند شناخته شده اند،
-
بر اساس دوره فصل موسمی جنوب غربی که معمولاً از ژوئن تا سپتامبر طول می کشد، از سال 1991 تا 2020، وقوع جغرافیایی فاخته ژاکوبن برای این ماه ها فیلتر شد. در این دوره، کل کشور بیش از 75 درصد دریافت می کند. بارندگی آن [ 98 ].
-
مجموعه ورودی دوم با استفاده از ماه های فصل موسمی شمال شرقی، یعنی اکتبر تا دسامبر فیلتر شد. این فصل موسمی همچنین به عنوان فصل پس از موسمی یا فصل موسمی زمستانی شناخته می شود که در آن کشور حدود 60 درصد از بارندگی سالانه خود را در نواحی ساحلی و حدود 40 درصد در مناطق داخلی دریافت می کند [ 99 ]. علاوه بر این، فصل بارانی از اکتبر شروع می شود و تا آوریل تا ژوئن در آفریقا ادامه می یابد، جایی که شرایط بیشتر برای اقامت آن مناسب است.
-
مجموعه سوم و آخر شامل داده های ماه های ژانویه تا مه است که به ترتیب دوره اواسط بارانی و پایان فصل بارانی در آفریقا و هند را نشان می دهد.
از این رو، مجموعه رویدادهای جغرافیایی ذکر شده در بالا با مجموعه داده های محیطی ترکیب شدند تا محدوده مناسب بالقوه، پارامترهای محیطی و تغییرات آب و هوایی تغییر یافته در سناریوهای مختلف تغییر آب و هوا را درک کنند.
3.3. داده های زیست محیطی
3.3.1. انتخاب متغیرهای محیطی
این بخش انتخاب دادههای محیطی را مورد بحث قرار میدهد که فرض میشود از نظر اکولوژیکی بر گونههای متحرک مانند پرندگان، به ویژه توزیع فاخته ژاکوبین تأثیر میگذارند. اینها شامل متغیرهای زیست اقلیم، حداقل دما، حداکثر دما، بارش، ارتفاع و باد با وضوح فضایی 2.5 قوس بر دقیقه از WorldClim است. برای شرایط اقلیمی کنونی، متغیرهای زیست اقلیمی، که برای سالهای 1970 تا 2000 بهطور میانگین محاسبه شدند، از WorldClim نسخه 2.1، آخرین نسخه دادههای آب و هوایی که در ژانویه 2020 راهاندازی شد [ 100 ] بهدست آمدند.]. طبق این مطالعه، مدلسازی برای سه دوره زمانی مختلف انجام شد. بنابراین، متغیرهای اقلیمی مانند بارش، باد، حداقل دما و حداکثر دما برای سه مجموعه داده شده گرفته و غربال شدند.
برای مجموعه های آینده، 19 متغیر زیست اقلیم ( جدول 1 ) برای آینده نزدیک (2021-2040) و آینده دور (2041-2060) پیش بینی نقشه های توزیع گونه ها در 2.5 قوس دقیقه از WorldClim [ 101 ] به دست آمد. برای سناریوهای اقلیمی آینده، دادههای اقلیمی CIMP مربوط به مدل جهانی آب و هوای CNRM-ESM2-1 [ 102 ] برای چهار مسیر اجتماعی-اقتصادی مشترک (SSP): 126، 245، 370 و 585 از پایگاه داده WorldClim که به صورت فضایی بود بهدست آمد. برای کاهش تعصب، مقیاس کوچک و کالیبره شده است.
پنجمین گزارش ارزیابی IPCC (هیئت بیندولتی تغییرات آب و هوا) 2013 (AR5) مدلهای آب و هوایی را از CMIP5 تولید کرد و ششمین گزارش ارزیابی IPCC 2021 (AR6) CMIP6 را با 10 مدل سیستم زمین (ESM) ارائه کرد [ 103]. CNRM-ESM2-1 یکی از ESMهایی است که شامل اجزای سیستم زمین تعاملی مانند ذرات معلق در هوا، شیمی جو و چرخه کربن زمین و اقیانوس است. در CMIP6، مقادیر کافی داده در Carbon Brief برای تجزیه و تحلیل سناریوهای انتشار آینده، مانند گرم شدن گذشته و آینده و حساسیت آب و هوا از CMIP5، گنجانده شد. IPCC AR5 چهار مسیر تمرکز نماینده (RCPs) را معرفی کرد که انتشار گازهای گلخانه ای آینده را در سناریوهای مختلف تغییرات آب و هوایی بررسی می کرد: RCP2.6، RCP4.5، RCP6.0 و RCP8.5. این سناریوها با سناریوهای مسیرهای اجتماعی و اقتصادی مشترک (SSP) در CMIP6–SSP1-2.6، SSP2-4.5، SSP3-7.0، SSP4-6.0 و SSP5-8.5 به روز شدند. SSP1 دنیای رشد و برابری متمرکز بر پایداری است. SSP2 به عنوان “وسط جاده” شناخته می شود، که در آن الگوهای تاریخی دنبال می شود. SSP3 درست در وسط دامنه نتایج پایه تولید شده توسط ESM ها قرار دارد، SSP4 دنیای خوش بینانه تری است که نمی تواند هیچ گونه سیاست اقلیمی را تعیین کند و SSP5 بدترین سناریو را به تصویر می کشد. این SSP ها می توانند عوامل جمعیت شناختی و اقتصادی و همچنین تأثیر انتخاب های اجتماعی بر انتشار گازهای گلخانه ای را بررسی کنند. در حالی که در RCPها، عوامل اجتماعی-اقتصادی در نظر گرفته نمیشوند، اما فقط مسیرهایی برای بررسی غلظت گازهای گلخانهای و میزان گرمایشی که میتواند تا پایان قرن رخ دهد تعیین میشود. در این مقاله از SSP های 1-2.6، 2-4.5، 3-7.0 و 5-8.5 استفاده شده است. این SSP ها می توانند عوامل جمعیت شناختی و اقتصادی و همچنین تأثیر انتخاب های اجتماعی بر انتشار گازهای گلخانه ای را بررسی کنند. در حالی که در RCPها، عوامل اجتماعی-اقتصادی در نظر گرفته نمیشوند، اما فقط مسیرهایی برای بررسی غلظت گازهای گلخانهای و میزان گرمایشی که میتواند تا پایان قرن رخ دهد تعیین میشود. در این مقاله از SSP های 1-2.6، 2-4.5، 3-7.0 و 5-8.5 استفاده شده است. این SSP ها می توانند عوامل جمعیت شناختی و اقتصادی و همچنین تأثیر انتخاب های اجتماعی بر انتشار گازهای گلخانه ای را بررسی کنند. در حالی که در RCPها، عوامل اجتماعی-اقتصادی در نظر گرفته نمیشوند، اما فقط مسیرهایی برای بررسی غلظت گازهای گلخانهای و میزان گرمایشی که میتواند تا پایان قرن رخ دهد تعیین میشود. در این مقاله از SSP های 1-2.6، 2-4.5، 3-7.0 و 5-8.5 استفاده شده است.
3.3.2. غربالگری متغیرهای محیطی
آزمون همبستگی بین متغیرهای محیطی برای هر یک از سه مجموعه فصلی برای حفظ متغیرهای مرتبط با محیط زیست در مناسب بودن گونه انجام شد. ضرایب همبستگی اسپیرمن [ 104 ] روی مجموعه متغیرها اعمال شد، که در آن، اگر متغیرها دارای همبستگی اسپیرمن <0.7 بودند، آن متغیرها همبستگی بالایی نداشتند. سپس، ضریب تورم واریانس (VIF) در نرم افزار R برای هر متغیر باقیمانده با استفاده از تابع vifcor بسته R usdm [ 105 ] محاسبه شد و متغیرهای محیطی که مقادیر VIF آنها > 3 است حذف شد، زیرا مقادیر VIF کوچکتر همبستگی پایینی دارند. مقادیر VIF حاصل <3 [ 106 ] بود]، و بنابراین، هیچ متغیر دیگری حذف نشد. این آزمون همبستگی بین متغیرهای محیطی برای هند و آفریقا به طور جداگانه انجام شد و سپس این متغیرهای محیطی غربالشده در مدل Maxent برای پیشبینی تناسب زیستگاه در مناطق مورد مطالعه فوق استفاده شد.
3.4. مدل توزیع فضایی با Maxent
مدلسازی Maxent مبتنی بر ML [ 107 ] محبوبترین و یک رویکرد مدلسازی مناسب زیستگاهی است که به خوبی تثبیت شده است [ 108 ، 109 ، 110 ، 111 ، 112 ، 113 ، 114 ، 115 ، 116 .] که توزیع های احتمالی را بر اساس وقوع گونه ها و متغیرهای محیطی پیش بینی می کند. مزیت استفاده از Maxent این است که از داده های حضوری استفاده می کند و یک نقشه پیش بینی در منطقه مورد مطالعه ارائه می دهد. این بر اساس اصل حداکثر آنتروپی کار می کند که توزیع احتمال زیستگاه گونه ها را بدون هیچ محدودیتی تخمین می زند و فرض می کند که هر ویژگی در توزیع تقریبی مقدار میانگین مشابه با وقوع گونه ها را دارد.
در این مطالعه، الگوریتم حداکثر آنتروپی با رخدادهای پرنده و متغیرهای پیشبین غربالشده برای پیشبینی زیستگاههای مناسب بالقوه و تحلیل اهمیت نسبی عوامل زیستاقلیمی مختلف هر نقطه برای فاخته ژاکوبن مدلسازی شد. این روش در هر سه مجموعه دوره زمانی اعمال شد، به طوری که تجزیه و تحلیل تناسب زیستگاه را می توان برای تأیید این باور که فاخته ژاکوبین منادی بادهای موسمی هند است و تجزیه و تحلیل آب و هوا و محدوده مناسب این پرنده در هند انجام داد. و همچنین در آفریقا، در دوره های انتخاب شده. آزمون jackknife برای تشخیص اهمیت متغیرهای محیطی استفاده شد. وقوع گونهها به ترتیب به عنوان دادههای آموزش (75 درصد از کل وقوع) و آزمون (25 درصد از کل وقوع) برای کالیبراسیون و ارزیابی مدلها تقسیم شدند. منحنی های پاسخ؛ چاقو و سایر ویژگی ها مانند خطی، درجه دوم، محصول، آستانه و لولا به عنوان پارامترهای واقعی در مدل مناسب زیستگاه تعیین شدند. سایر پارامترهای مدل به شرح زیر استفاده شد:
-
“replicates = 10” به مدل در مورد تعداد تکرارهایی که مدل برای اعتبارسنجی متقابل، بوت استرپ یا انجام نمونه برداری با اجراهای جایگزین اجرا می کند، می گوید.
-
“lq2lqptthreshold = 80” تعداد نمونه هایی است که در آن ویژگی های محصول و آستانه شروع به استفاده می کنند.
-
“l2lqthreshold = 10” تعداد نمونه هایی است که در آن ویژگی های درجه دوم شروع به استفاده می کنند و
-
“آستانه لولا = 15” تعداد نمونه هایی است که در آن ویژگی های لولا شروع به استفاده می کنند.
سپس عملکرد پیشبینی مدل تولید شده با محاسبه مساحت زیر منحنی اپراتور گیرنده (AUC) نمودار مشخصه عملکرد گیرنده (ROC) ارزیابی شد که بین 0 (بدون تبعیض) تا 1 (تبعیض کامل) است [ 116 ].]. فرآیند ارزیابی عملکرد پیشبینی مدل با استفاده از AUC شامل فرآیند تعیین آستانههایی برای پیشبینی مدل با تولید سطوح مختلف نرخهای مثبت کاذب و سپس ارزیابی نرخ مثبت واقعی به عنوان تابع نرخ مثبت کاذب است. در اینجا، نرخ مثبت کاذب به پیشبینی حضور برای مکانهایی که گونه غایب است، اشاره دارد و نرخ مثبت واقعی، پیشبینی موفق حضور است. محدوده AUC از 0.7 تا 0.8 قابل قبول است، 0.8 تا 0.9 عالی است و بالای 0.9 عملکرد فوق العاده ای است [ 117 ]. متغیرهای محیطی غالب در تعیین پراکنش احتمالی گونهها از طریق آزمون jackknife (همچنین به نام ترک یکاوت) مورد ارزیابی قرار گرفتند که به جایگشت در برابر متغیرهای محیطی اهمیت میدهد.110 ]. منحنیهای پاسخ گونهها توسط این مدل برای بررسی چگونگی واکنش احتمال وقوع گونهها به تغییرات در شرایط متغیر محیطی ایجاد شد.
سپس، تغییرات اقلیمی آینده (2021-2040 و 2041-2060) نیز برای برآورد چگونگی واکنش گونه ها به تغییرات در سیستم های اکولوژیکی، مدل سازی شد، زیرا زیستگاه های مطلوب آنها تحت سناریوهای مختلف تغییر آب و هوا (به عنوان مثال، SSP1-2.6، SSP2) تغییر خواهد کرد. -4.5، SSP3-7.0 و SSP5-8.5).
نقشههای تناسب زیستگاه پیشبینیشده سپس به کلاسهای مناسبی طبقهبندی شدند که نشاندهنده محدودیتهای آستانهای بود که زیستگاههای نامناسب و مناسب را متمایز میکرد. طبقات بازطبقه بندی مناسب زیستگاه عبارت بودند از: شرایط نامناسب با آستانه پایین تر و شرایط مناسب که در ادامه به سه طبقه کم، متوسط و بسیار مناسب دسته بندی شدند. این آستانه با شناسایی مناطقی که حداقل مناسب با مناطقی که گونه ثبت شده است، به تفسیر اهمیت اکولوژیکی کمک کرد.
4. نتایج
4.1. انتخاب متغیرهای محیطی
برای تشخیص همبستگی بین متغیرهای محیطی، آستانه ضریب همبستگی اسپیرمن 0.7 تعیین شد و سپس تابع vifcor انجام شد. از ارزیابی بصری، برخی از متغیرها یک همبستگی متقابل را نشان دادند که اگر مقادیر vif آنها کمتر از 3 باشد، حذف شد. سپس مجموعه نهایی انتخاب شده از متغیرها برای پیش بینی مناسب بودن فاخته ژاکوبین، ارائه شده در جدول 2 استفاده شد.
4.2. نتایج ارزیابی عملکرد مدل Maxent
پس از اجرای مدل Maxent بر روی وقوع گونه ها و متغیرهای محیطی، دقت پیش بینی آن با استفاده از نمودارهای AUC مورد ارزیابی قرار گرفت. همانطور که در جدول 2 نشان داده شده است ، متغیرهای محیطی برای هند و آفریقا به طور جداگانه در سه دوره زمانی مختلف انتخاب شدند. بنابراین، مدل Maxent با تفکیک گونههای رخدادهای هند و آفریقا در سه دوره زمانی مختلف اجرا شد. همچنین حداقل دما، حداکثر دما و بارندگی مورد استفاده در مدل Maxent با توجه به این سه بازه زمانی گرفته شد.
مقادیر AUC برای مدل پیشبینی تناسب فاختههای Pied ارائهشده در جدول 3 نشان میدهد که پیشبینی مدل بسیار خوب بود، به طوری که میتواند به طور موثر توزیع گونهها را تحت سناریوهای آب و هوایی فعلی و آینده پیشبینی کند.
4.3. اهمیت و سهم متغیر
جدول 4 و جدول 5 تخمین اکتشافی سهم درصد و اهمیت جایگشت متغیرهای محیطی مورد استفاده در مدل Maxent را برای سه دوره زمانی مختلف با دادههای وقوع گونهها از هند و آفریقا نشان میدهند. این دو جدول به ما کمک کرد تا تأثیرگذارترین متغیرهای محیطی را که نقش مهمی در مناسب بودن زیستگاه فاخته ژاکوبن در هند و آفریقا داشتند، تفسیر کنیم. در جدول 4 مشاهده شده است که bio2، bio3، bio14، bio15، bio18، bio19 و باد برای هر سه دوره زمانی در هند مشترک هستند، در حالی که در آفریقا ( جدول 5)) bio 14 و bio 19 در هر سه دوره زمانی مشترک هستند. در هند، باد و بارش نقش کوچکی دارند، در حالی که در آفریقا، باد و ارتفاع سهم عمدهای در مدلسازی مناسب دارند. بنابراین، این بخش به این نتیجه رسید که متغیرهای محیطی مربوط به بارش نقش بسزایی در پراکنش فاخته ژاکوبن ایفا میکنند و اساساً در زیستگاههای بالقوه مناسب آن مورد نیاز هستند.
4.4. نقشه تناسب زیستگاه پیش بینی شده فاخته ژاکوبین
با استفاده از عوامل زیست اقلیمی موثر در پراکنش گونهها، پیشبینی تناسب زیستگاه تحت سناریوهای اقلیمی فعلی و آتی برای برآورد تغییرات در سیستمهای اکولوژیکی و چگونگی واکنش گونهها به تغییرات در تغییرات مختلف آب و هوایی انجام شد. این بخش ویژگی های فضایی داده های توزیع مورد استفاده را مورد بحث قرار می دهد، به عنوان مثال، فصل های موسمی جنوب غربی و شمال شرقی هند و فصل بارانی آفریقا، به ویژه در جنوب آفریقا.
4.4.1. تناسب زیستگاه فعلی
ژوئن-سپتامبر
ماههای ژوئن تا سپتامبر به عنوان دوره بارانهای موسمی جنوب غربی هند شناخته میشوند، که در آن تمام هند بیش از 75 درصد از بارندگی خود را دریافت میکند [ 118 ]، در حالی که این ماهها فصل خشک را در آفریقای مرکزی، جنوبی و شرقی به ارمغان میآورند. بنابراین، رخدادهای ثبت شده در ماه های ژوئن تا سپتامبر (به طور جداگانه برای هند و آفریقا) با متغیرهای محیطی غربال شده به مدل Maxent ارائه شد که منجر به ایجاد متغیرهای زیست اقلیم غالب و همچنین پیش بینی مناسب بودن زیستگاه فعلی این پرنده شد. پس از ارزیابی عملکرد مدل، خروجی برای پیش بینی مناسب بودن زیستگاه آینده فاخته ژاکوبین تحت سناریوهای مختلف تغییر آب و هوا مورد استفاده قرار گرفت.
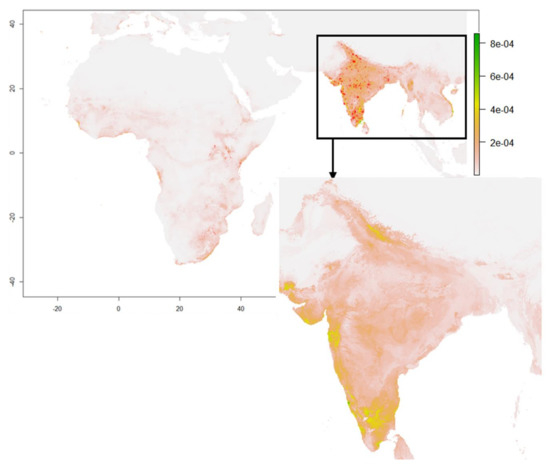
نقشه تناسب زیستگاه گونه ها در شکل 3 نشان داده شده استنشان می دهد که مناطق پوشیده از رنگ خاکستری برای این پرنده مناسب نیست، در حالی که رنگ های زرد و قهوه ای به ترتیب نشان دهنده مناسب بودن زیستگاه خوب و کم گونه برای این پرنده است. وقوع گونه ها با نقاط قرمز رنگ بر روی نقشه ترسیم شده است و محدوده مناسب زیستگاه را می توان از مقیاس احتمال که محل سکونت پرنده را نشان می دهد، مشاهده کرد. بر این اساس، آفریقا در طول دوره ژوئن-سپتامبر به دلیل فصل خشک آن، مناسب بودن بسیار پایینی دارد که ممکن است آب و هوای مناسبی برای مناسب بودن فاخته های پایی نباشد. با این حال، مدل مناسب بودن خوب و بالای این پرندگان را در کل هند در مقایسه با آفریقا پیش بینی کرد، زیرا هند در آن زمان 75 درصد بارندگی خود را دریافت می کند و بنابراین، این می تواند یکی از دلایل اصلی مهاجرت جزئی آنها به هند باشد. تا بتوانند عوامل اقلیمی مناسب خود را بدست آورند.شکل 3 بر روی رخدادهای 1991 تا 2020 و متغیرهای آب و هوایی (tmin، tmax، prec و wind) ماه های ژوئن تا سپتامبر محاسبه شده است.
در هند، عمدهترین پیشبینیهای تناسب در سناریوی کنونی را میتوان در شمال، غرب و جنوب هند مشاهده کرد، اما شرق، مرکز و شمال شرق هند پیشبینیهای بسیار کم یا بدون مناسب بودن را نشان دادهاند. این مدل میانگین محدوده مناسب فاخته ژاکوبین را در ایالتهای هندی زیر – اوتاراکند و اوتار پرادش و در مادیا پرادش در هند مرکزی پیشبینی کرد. دلیل مهاجرت آن به این مناطق از هند می تواند به دلیل مرطوب ترین ویژگی ها به دلیل بیشترین میزان بارندگی و حداکثر تعداد رودخانه ها باشد. محدوده مناسبی خوبی در غرب هند، مانند گجرات (محلهای نزدیک به خلیج کاچچا و خلیج خمبات) و در ماهاراشترا (محلهای احاطه شده توسط دریای عرب) و همچنین در جنوب هند، مانند گات های غربی بیشترین تناسب این پرنده در چند منطقه از جنوب هند مانند جنوب تامیل نادو که عبارتند از Rameswaram، Dhanushkodi و Thoothukodi پیش بینی شده است. بنابراین، این احتمال وجود دارد که این پرنده دوست دارد در مرطوب ترین مکان ها که بیشترین میزان بارندگی را دریافت می کنند، بماند. در آفریقا، هیچ گونه مناسب زیستگاهی برای فاخته ژاکوبین پیش بینی نشده بود، زیرا این دوره زمانی فصل خشک است که ممکن است باعث مهاجرت فاخته ژاکوبین به هند شود.
تغییر در آب و هوا، به ویژه الگوهای موسمی جنوب غربی هند، با توجه به فاخته ژاکوبین در 30 سال گذشته با جدا کردن مجموعه دادهها به دو زیر مجموعه – 1991-2005 و 2006-2020 تجزیه و تحلیل شد. برای این منظور از متغیرهای اقلیمی برای این زیرمجموعه های سالانه استفاده شد که نتایج آنها به ترتیب در شکل 4 و شکل 5 نشان داده شده است. در شکل 4 ، هیچ تناسبی در آفریقا در سال های 1991-2006 در طول ژوئن تا سپتامبر پیش بینی نشده است، زیرا آب و هوای خشک آن برای پرنده نامناسب است، اما، در هند، تناسب زیستگاه مناسب را می توان در بخش های شرقی و شمالی مشاهده کرد. به دلیل مهاجرت پرنده به هند در جستجوی آب و هوای مرطوب، بخشهای جنوبی و غربی آن نه. زمانی که تناسب نتایج ازشکل 4 با شکل 5 مقایسه شده است، کاهشی در بارندگی موسمی هند در شمال و شرق طی 15 سال گذشته در ژوئن تا سپتامبر مشاهده شده است. علاوه بر این، این مطالعه در مورد تغییرات آب و هوایی کاملاً به مشاهده فاخته ژاکوبن وابسته بود، بنابراین یکی از دلایل مناسب بودن کمتر در جنوب و غرب هند، مشاهده کمتر پرنده به دلیل عدم آگاهی از جمع سپاری است. با توجه به سوابق جستجوی Google Trends از سال 2004 تا کنون ( شکل 6 )، علاقه عمومی به اصطلاح “انبوه سپاری” در هند در سال 2007 آغاز شد.
اکتبر-دسامبر و ژانویه-مه
ماه های اکتبر، نوامبر و دسامبر به موسمی شمال شرقی یا بادهای موسمی زمستانی در هند معروف هستند، زیرا جهت آن از شمال شرقی به جنوب غربی هند و آغاز فصل بارانی و مرطوب در آفریقا است که معمولاً تا آوریل ادامه دارد. به این ترتیب، این مجموعه ماه ها شامل موسمی های دو کشور مختلف است. بنابراین، نتایج مورد بحث در این بخش به دو بخش فرعی تقسیم می شود: یکی برای اکتبر تا دسامبر و دیگری برای ژانویه تا مه.
فصل بادهای موسمی شمال شرقی از اکتبر تا دسامبر در هند باران را به آندرا پرادش عمدتاً در مناطق ساحلی کرالا، پودوچری، رایالاسیما، کارناتاکای جنوبی و تامیل نادو میبارد. در مقایسه با بادهای موسمی جنوب غربی، این دوره موسمی تنها 11 درصد از بارندگی سالانه در هند را به خود اختصاص می دهد، اما در تامیل نادو، این فصل تقریباً نیمی از بارندگی سالانه خود را دارد. نقشه تناسب زیستگاه ارائه شده در شکل 7نشان می دهد که در هند، بالاترین تناسب زیستگاه فاخته ژاکوبین در تامیل نادو، خوب در آندرا پرادش و کارناتاکا و کم در کرالا پیش بینی شده است. با توجه به پیشبینیهای مدل در مورد مناسب بودن زیستگاه پرنده، که با الگوهای بادهای موسمی شمال شرقی (یعنی در ماههای اکتبر، نوامبر و دسامبر) همبستگی دارد، میتوان آن را به این باور یا واقعیت مرتبط دانست که حرکات پرنده کاملاً با موسمی مرتبط است. باران های دوره موسمی شمال شرقی.
پس از بررسی و کاوش ارتباط ورود موسمی هند با مشاهده پرنده، متعاقباً، همان مطالعه برای بررسی عوامل عمده اقلیمی که می تواند علت سفر بازگشت پرنده به آفریقا باشد، انجام شد. شکل 7 و شکل 8نشان می دهد که وقتی فصل بارانی و مرطوب در ماه اکتبر شروع می شود، فاخته های پایید، ساکنان آفریقا، ممکن است به سرزمین مادری خود بازگردند و تا آوریل در آنجا اقامت کنند. بنابراین مشاهده فاخته ژاکوبن در استان های آفریقا به جز کیپ غربی مشاهده شد. با این حال، بالاترین تناسب (رنگ سبز) در مناطق ساحلی کیپ شرقی و استان کوازولو-ناتال پیشبینی میشود. بنابراین، تحقیق در مورد مناسب بودن زیستگاه فاخته ژاکوبین ارتباط بین مشاهده فاخته ژاکوبین و رسیدن فصل باران های موسمی در هند را ثابت کرد و همچنین داستان های اجدادی یا باورهای سنتی را به وجود آورد که این فاخته های ژاکوبنی توانایی جادویی احضار را دارند. هر جا که بروند باران ببارند، مانند سراسر شبه قاره هند و همچنین در آفریقا.
4.4.2. مناسب بودن آینده پیش بینی شده تحت سناریوهای مختلف تغییر آب و هوا
این بخش مربوط به مدلسازی تناسب زیستگاه آتی فاخته ژاکوبین با استفاده از مدل مکسنت موجود، دادههای رخدادهای موجود و لایههای محیطی موجود و سپس پیشبینی آنها با متغیرهای زیستمحیطی آتی سالهای 2030 و 2050 میباشد. علاوه بر این، افزایش احتمالی و یا کاهش زیستگاه های مناسب و نامناسب در سال های جاری و آینده نیز برای سایت های اشغال شده توسط گونه تخمین زده شد که می توان از آن در مطالعات مختلف تغییرات اقلیمی بعد استفاده کرد.
ژوئن-سپتامبر
سپس نقشه های تناسب حاصل با استفاده از متغیرهای محیطی انتخاب شده که در شکل 9 آورده شده است، تولید شد . شکل نشان می دهد که شرایط مناسب زیستگاه احتمالی فاخته ژاکوبین در SSP 2.6، 4.5 و 7.0 سال 2030 نسبتاً متوسط است. در مقایسه با پیش بینی های فعلی در شکل 5 ، کاهش در سناریوهای اقلیمی مختلف سال های آینده، به ویژه مشاهده می شود. در SSP 8.5، که در آن پیکسل های مناسب ناپدید شدند. این ممکن است به دلیل تخمین انتشار بیشتر CO 2 و افزایش گرمایش جهانی باشد. جدول 6 نشان می دهد که در مناطق مناسب کاهش و در مناطق نامناسب نسبت به مناطق فعلی افزایش یافته است.
اکتبر-دسامبر و ژانویه-مه
این بخش در مورد چگونگی تغییر توزیع زیستگاه های بالقوه تحت سناریوهای مختلف تغییرات آب و هوایی بحث می کند. با توجه به پیشبینیهای آینده مدل برای ماههای اکتبر-دسامبر در هند ( شکل 10 ) و آفریقا ( شکل 11 )، مناسب بودن زیستگاه فاخته ژاکوبین به شدت به سناریوی فعلی ( شکل 7 ) تحت SSPs 2.6 و 4.5 مربوط میشود. 2030. با این حال، شرایط مناسب در سناریوهای مختلف مانند 7.0 و 8.5 سال 2030 و در سال 2050، تحت تمام سناریوهای اقلیمی، احتمال وقوع فاخته ژاکوبین در مقایسه با سناریو فعلی بسیار کم پیشبینی میشود. این را می توان در جدول 7 و جدول 8 مشاهده کردکه نشان دهنده افزایش نامناسب و کاهش مناطق مناسب فاخته ژاکوبن در هند و آفریقا است.
سایر پیشبینیهای آینده مناسب برای ماههای ژانویه تا مه 2030 و 2050 از طریق مدل Maxent به ترتیب در شکل 12 و شکل 13 برای هند و آفریقا نشان داده شده است، که این احتمال را فراهم میکند که تناسب پرنده در سال 2050 تحت همه سناریوهای اقلیمی کم شود. . چنین کاهشی در تناسب زیستگاه پرنده در این ماه ها نشان می دهد که در آینده، هند و همچنین آفریقای جنوبی ممکن است باران کمتر و خشکی بیشتری داشته باشند که منجر به کاهش تناسب فاخته ژاکوبین در هند خواهد شد ( جدول 9 ) و آفریقا ( جدول 10 ). اگرچه ممکن است بین رویکردهای مختلف مدلسازی آب و هوا ناسازگاری وجود داشته باشد [ 119]، استراتژی ارزیابی تناسب فعلی و پیشبینی تغییرات آتی در پراکنش گونههای متنوع، که تحت تأثیر الگوهای اقلیمی مختلف قرار دارند، همچنان به عنوان یک حوزه تحقیقاتی مهم شناخته میشود.
5. بحث و گفتگو
مطالعه ارائه شده در اینجا مناسب بودن زیستگاه برای فاخته ژاکوبین را در فصول مختلف، با اشاره خاص به هند، با استفاده از وقوع گونه (1991-2020) در مدل Maxent مبتنی بر ML با متغیرهای محیطی تجزیه و تحلیل کرد. وقایع از پایگاه داده GBIF، رصدخانه ای متشکل از داده های مؤسسات عمومی، به عنوان مثال، موزه ها، و مشاهدات شهروندان، به دست آمد. عملکرد پیش بینی مدل Maxent به مقادیر AUC بالاتری دست می یابد که نشان دهنده عالی و دقیق بودن این مدل است. نتایج به دست آمده با استفاده از روش Maxent برای پیش بینی مناسب بودن بالقوه فاخته ژاکوبین در هر سه فصل هند یعنی ژوئن-سپتامبر (موسمی جنوب غربی)، اکتبر-دسامبر (شمال موسمی شمال شرقی) و ژانویه-مه (زمستان) متفاوت است. و تابستان). متغیرهای مهم محیطی موثر بر تناسب زیستگاه آن عبارتند از: بارش خشک ترین ماه، فصلی بودن بارش، بارش گرم ترین فصل، میانگین دمای مرطوب ترین فصل و باد. پیشبینیهای مدل نشان داد که مناسب بودن گونهها از همان الگوی هر دو فصل بارانهای موسمی هند، یعنی جنوب غربی و شمال شرقی پیروی میکند. بنابراین، بر اساس نتایج، مهاجرت پرنده را می توان با بادهای موسمی در مناطق ارزیابی شده – هند و آفریقا – مرتبط کرد. با این وجود، به منظور بررسی فصل بادهای موسمی جنوب غربی هند، مجموعه دادهها به دو زیر مجموعه 1991-2005 و 2006-2020 تقسیم شدند و سپس، مدل Maxent با دادههای محیطی اجرا شد. از نتایج، مشاهده اینکه الگوهای موسمی در سال 1991 در چند منطقه شمال و شرق هند در طول دوره ژوئن تا سپتامبر شروع به کاهش کردند که ممکن است به دلیل فعالیت های انسانی، جنگل زدایی و غیره باشد، به طرز شگفت آوری جالب بود. شرایط آب و هوایی همیشه برای اقامت این پرنده از مهر ماه و تا فروردین ماه مناسب است. هنگامی که فصل بارانی و مرطوب در آفریقا به پایان می رسد، پرندگان شروع به مهاجرت به نقاط مختلف جهان می کنند، جایی که شرایط آب و هوایی مطلوب تری دارند. مناسب بودن آینده پرنده فاخته Pied در اینجا با مجموعه کاملی از شرایط آب و هوایی تحت چهار سناریو (SSPs) مدلسازی شد: 2.6، 4.5، 7.0 و 8.5 برای سال 2030 (میانگین برای 2021-2040) و 2050 (میانگین برای 2060-222) ، با استفاده از نتایج تناسب فعلی و متغیرهای زیست اقلیم پیش بینی شده آن. طبق پیش بینی های آینده انجام شده در این مطالعه، توزیع آب و هوایی بالقوه مناسب در آینده (2050 < 2030 < فعلی) تحت سناریوهای مختلف تغییر آب و هوا کاهش می یابد، که نشان می دهد ممکن است تغییری در فصل باران های موسمی در هند نیز وجود داشته باشد. مانند آفریقا، که منجر به تناسب کمتر برای فاخته ژاکوبن می شود. چنین پیوند مستقیمی از این پرنده با فصل باران های موسمی به تجزیه و تحلیل انتقادی فعالیت های احتمالی تغییرات آب و هوایی و اینکه کدام متغیرهای محیطی برای مناسب بودن آن و حمایت از جنبش های مهاجرتی آن نقش موثری دارند، کمک می کند. این نشان می دهد که ممکن است تغییری در فصل باران های موسمی در هند و همچنین در آفریقا رخ دهد که منجر به تناسب کمتر برای فاخته ژاکوبین می شود. چنین پیوند مستقیمی از این پرنده با فصل باران های موسمی به تجزیه و تحلیل انتقادی فعالیت های احتمالی تغییرات آب و هوایی و اینکه کدام متغیرهای محیطی برای مناسب بودن آن و حمایت از جنبش های مهاجرتی آن نقش موثری دارند، کمک می کند. این نشان می دهد که ممکن است تغییری در فصل باران های موسمی در هند و همچنین در آفریقا رخ دهد که منجر به تناسب کمتر برای فاخته ژاکوبین می شود. چنین پیوند مستقیمی از این پرنده با فصل باران های موسمی به تجزیه و تحلیل انتقادی فعالیت های احتمالی تغییرات آب و هوایی و اینکه کدام متغیرهای محیطی برای مناسب بودن آن و حمایت از جنبش های مهاجرتی آن نقش موثری دارند، کمک می کند.
6. نتیجه گیری
این مطالعه به این نتیجه رسید که سیستمهای اکولوژیکی با توجه به تغییرات اقلیمی تغییر خواهند کرد و زیستگاههای مطلوب گونهها تحت سناریوهای مختلف تغییر اقلیم تغییر خواهند کرد. مطالعه حاضر نمونهای از مدلسازی یا پیشبینی این تغییرات را با استفاده از مشاهدات شهروندان نشان میدهد که مجموعه دادههای مورد نیاز یا مشاهدات را برای اعمال مدلهای ML قوی ارائه میدهد. بنابراین، استفاده از روشهای شهروندی برای امکانپذیر کردن چنین تحلیلی ضروری بود. مدل سازی مناسب بودن آینده با استفاده از مجموعه داده های آینده CMIP6 نشان داد که بارش و مرطوب ترین اقلیم ممکن است کاهش یابد در حالی که آب و هوای گرم و خشک ممکن است افزایش یابد.
مرطوب ترین فصل و بارندگی عناصر اصلی در پراکنش فاخته های ژاکوبین هستند و برنامه های مشترک مختلفی برای حفظ تناسب پرندگان مهاجر مختلف مانند فاخته ژاکوبین در چنین گرم شدن پتانسیل متغیر و غیرمنتظره زمین مورد نیاز است. با این حال، چنین تغییرات پیشبینیشده تنها بر اساس عوامل اقلیمی است و لزوماً به توزیع کاربری اراضی تحت اشغال انسان مانند سکونتگاههای شهری و توانایی پراکندگی مرتبط نیست.
منابع
- Araújo، MB; Guisan، A. پنج چالش (یا بیشتر) برای مدلسازی توزیع گونهها. J. Biogeogr. 2006 ، 33 ، 1677-1688. [ Google Scholar ] [ CrossRef ]
- ماتیوپولوس، جی. فیبرگ، جی. آرتس، GA؛ Beyer, HL; مورالس، جی.ام. Haydon، DT برقراری ارتباط بین انتخاب زیستگاه و پویایی جمعیت حیوانات. Ecol. مونوگر. 2015 ، 85 ، 413-436. [ Google Scholar ] [ CrossRef ]
- هیرزل، ق. لی لی، جی. هلفر، وی. راندین، CF; Guisan، A. ارزیابی توانایی مدلهای تناسب زیستگاه برای پیشبینی حضور گونهها. Ecol. مدل. 2006 ، 199 ، 142-152. [ Google Scholar ] [ CrossRef ]
- فیلیپس، اس جی. اندرسون، آر.پی. دودیک، م. Schapire، RE; بلر، من باز کردن جعبه سیاه: نسخه منبع باز Maxent. اکوگرافی 2017 ، 40 ، 887-893. [ Google Scholar ] [ CrossRef ]
- یان، اچ. فنگ، ال. ژائو، ی. وو، دی. زو، سی. پیشبینی توزیع فضایی Alternanthera philoxeroides در چین بر اساس ArcGIS و MaxEnt. گلوب. Ecol. حفظ کنید. 2020 ، 21 ، e00856. [ Google Scholar ] [ CrossRef ]
- روندینی، سی. استوارت، اس. Boitani، L. مدل های مناسب زیستگاه و کمبود در برنامه ریزی حفاظتی برای مهره داران آفریقایی. حفظ کنید. Biol. 2005 ، 19 ، 1488-1497. [ Google Scholar ] [ CrossRef ]
- Tikkanen، O.-P. هاینونن، تی. کوکی، جی. مدلهای مناسب بودن زیستگاه ماترو، J. گونههای جنگلهای شمالی فهرست قرمز ساپروکسیلیک در مدیریت طولانیمدت ماتریس: اقدامات مقرونبهصرفه برای حفاظت از چند گونه. Biol. حفظ کنید. 2007 ، 140 ، 359-372. [ Google Scholar ] [ CrossRef ]
- رمیا، ک. راماچاندران، ا. Jayakumar، S. پیش بینی توزیع زیستگاه مناسب فعلی و آینده Myristica dactyloides Gaertn. با استفاده از مدل MaxEnt در گات شرقی، هند. Ecol. مهندس 2015 ، 82 ، 184-188. [ Google Scholar ] [ CrossRef ]
- Millar, CS; Blouin-Demers، G. مدل سازی مناسب زیستگاه برای گونه های در معرض خطر به الگوریتم و مقیاس حساس است: مطالعه موردی لاک پشت Blanding، Emydoidea blandingii، در انتاریو، کانادا. جی. نات. حفظ کنید. 2012 ، 20 ، 18-29. [ Google Scholar ] [ CrossRef ]
- لطیف، QS; ساب، VA; دادلی، جی جی; مارکوس، ا. Mellen-McLean، K. توسعه و ارزیابی مدل های مناسب زیستگاه برای لانه سازی دارکوب سر سفید (Dryobates albolarvatus) در جنگل سوخته. PLoS ONE 2020 , 15 , e0233043. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- تیتنسور، DP; باکو، آر. بروین، PE; کلارک، ام آر؛ کنسالوی، ام. هال اسپنسر، جی. رودن، آ. شلاچر، تی. سهام، KI; راجرز، AD پیش بینی مناسب بودن زیستگاه جهانی برای مرجان های سنگی در کوه های دریایی. J. Biogeogr. 2009 ، 36 ، 1111-1128. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- شارما، اس. آروناچلام، ک. بهاوسار، د. کالا، آر. مدلسازی مناسب بودن زیستگاه Perilla frutescens با MaxEnt در اوتاراکند – یک رویکرد حفاظتی. J. Appl. Res. پزشکی معطر. گیاهان 2018 ، 10 ، 99-105. [ Google Scholar ] [ CrossRef ]
- احمد، SE; مک اینرنی، جی. اوهارا، ک. هارپر، آر. سالیدو، ال. اموت، اس. Joppa، دانشمندان LN و نرم افزار-بررسی جامعه مدل سازی توزیع گونه ها. غواصان. توزیع کنید. 2015 ، 21 ، 258-267. [ Google Scholar ] [ CrossRef ]
- الیت، جی. گراهام، CH; اندرسون، آر.پی. دودیک، م. فریر، اس. گیسان، ع. Hijmans، RJ; Huettmann، F. Leathwick، JR; لمان، ا. و همکاران روشهای جدید پیشبینی توزیع گونهها را از دادههای وقوع بهبود میبخشند. اکوگرافی 2006 ، 29 ، 129-151. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- هرناندز، PA; گراهام، CH; استاد، LL; آلبرت، DL اثر اندازه نمونه و ویژگیهای گونه بر عملکرد روشهای مدلسازی توزیع گونههای مختلف. اکوگرافی 2006 ، 29 ، 773-785. [ Google Scholar ] [ CrossRef ]
- کادمون، ر. فاربر، او. Danin، A. تجزیه و تحلیل سیستماتیک عوامل مؤثر بر عملکرد مدل های پوشش اقلیمی. Ecol. Appl. 2003 ، 13 ، 853-867. [ Google Scholar ] [ CrossRef ]
- اسکیدمور، AK; گولد، آ. Walker, P. طبقه بندی توزیع زیستگاه کانگوروها با استفاده از سه مدل GIS. بین المللی جی. جئوگر. Inf. سیستم 1996 ، 10 ، 441-454. [ Google Scholar ] [ CrossRef ]
- استاکول، DR; پترسون، AT اثرات حجم نمونه بر دقت مدلهای توزیع گونهها. Ecol. مدل. 2002 ، 148 ، 1-13. [ Google Scholar ] [ CrossRef ]
- ویز، ام اس; Hijmans، RJ; پترسون، AT; گراهام، CH; گیسان، ع. کارگروه توزیع گونه های پیش بینی NCEAS. اثرات حجم نمونه بر عملکرد مدل های توزیع گونه ها. غواصان. توزیع کنید. 2008 ، 14 ، 763-773. [ Google Scholar ] [ CrossRef ]
- چپمن، AD; Grafton, O. Guide to Best Practices for Generalising Sensitive/Primary species Occurrence-Data, Version 1.0 ; تسهیلات جهانی اطلاعات تنوع زیستی: کپنهاگ، دانمارک، 2008; جلد 27. [ Google Scholar ] [ CrossRef ]
- گراهام، CH; فریر، اس. هوتمن، اف. موریتز، سی. پترسون، AT تحولات جدید در انفورماتیک مبتنی بر موزه و کاربردها در تجزیه و تحلیل تنوع زیستی. Trends Ecol. تکامل. 2004 ، 19 ، 497-503. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- ویزورک، جی. گوا، کیو. Hijmans, R. روش نقطه-شعاع برای ارجاع جغرافیایی توصیفات محل و محاسبه عدم قطعیت مرتبط. بین المللی جی. جئوگر. Inf. علمی 2004 ، 18 ، 745-767. [ Google Scholar ] [ CrossRef ]
- فیلی، کی جی. سیلمن، MR مدلسازی پاسخ گونههای گیاهی آند و آمازون به تغییرات آب و هوایی: اثرات خطاهای ارجاع جغرافیایی و اهمیت فیلتر کردن دادهها. J. Biogeogr. 2010 ، 37 ، 733-740. [ Google Scholar ] [ CrossRef ]
- گوا، کیو. لیو، ی. Wieczorek، J. ارجاع جغرافیایی توصیفات محلی و محاسبه عدم قطعیت مرتبط با استفاده از یک رویکرد احتمالی. بین المللی جی. جئوگر. Inf. علمی 2008 ، 22 ، 1067-1090. [ Google Scholar ] [ CrossRef ]
- گراهام، CH; الیت، جی. Hijmans، RJ; گیسان، ع. پترسون، AT; لویزل، بی. کارگروه توزیع گونه های پیش بینی Nceas. تأثیر خطاهای مکانی در دادههای وقوع گونهها که در مدلهای توزیع استفاده میشوند. J. Appl. Ecol. 2007 ، 45 ، 239-247. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- دیکینسون، جی ال. شرک، ج. Bonter، DN; بانی، آر. کرین، RL; مارتین، جی. فیلیپس، سل؛ Purcell, K. وضعیت فعلی علم شهروندی به عنوان ابزاری برای تحقیقات زیست محیطی و مشارکت عمومی. جلو. Ecol. محیط زیست 2012 ، 10 ، 291-297. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- الیت، جی. مدلهای توزیع گونههای Leathwick، JR: توضیح و پیشبینی بومشناختی در فضا و زمان. آنو. کشیش اکول. تکامل. سیستم 2009 ، 40 ، 677-697. [ Google Scholar ] [ CrossRef ]
- فرانکلین، جی. نقشه برداری گونه ها: استنتاج و پیش بینی فضایی . انتشارات دانشگاه کمبریج: کمبریج، بریتانیا، 2010. [ Google Scholar ]
- Pocock، MJO; تودل، جی سی. ساویج، جی. رابینسون، ال. روی، HE تنوع و تکامل علم شهروندی اکولوژیکی و زیست محیطی. PLoS ONE 2017 , 12 , e0172579. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- تولوچ، هوش مصنوعی؛ پوسینگام، اچ پی؛ جوزف، LN; سابو، جی. مارتین، TG درک پتانسیل کامل برنامه های نظارت بر علم شهروندی. Biol. حفظ کنید. 2013 ، 165 ، 128-138. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- سینگ، پی. ساران، اس. مدلسازی حداکثر آنتروپی با استفاده از علم شهروندی: مورد استفاده در فاخته ژاکوبین به عنوان شاخصی از موسمی هند. ISPRS Ann. فتوگرام از راه دور. حس اسپات. Inf. علمی 2020 ، 5 ، 23-30. [ Google Scholar ] [ CrossRef ]
- وایت لاو، جی. وان، اچ. کریگ، بی. اتکینسون، دی. ایجاد شبکه نظارت بر جامعه کانادا. محیط زیست نظارت کنید. ارزیابی کنید. 2003 ، 88 ، 409-418. [ Google Scholar ] [ CrossRef ]
- کوپر، سی بی; دیکینسون، جی. فیلیپس، تی. Bonney, R. Citizen Science به عنوان ابزاری برای حفاظت در اکوسیستم های مسکونی. Ecol. Soc. 2007 ، 12 ، 11. [ Google Scholar ] [ CrossRef ]
- هاو، جی. ظهور جمع سپاری. مگ سیمی 2006 ، 14 ، 1-4. [ Google Scholar ]
- ایروین، آ. ساخت شهروند علمی: علم و دموکراسی در علوم زیستی. پیکربندی مجدد طبیعت 2017 ، 10 ، 281-310. [ Google Scholar ] [ CrossRef ]
- بورک، جی. استرین، دی. هانسن، ام. پارکر، ای. راماناتان، ن. ردی، اس. Srivastava، MB حس مشارکتی. UCLA: مرکز سنجش شبکه جاسازی شده 2006. در دسترس آنلاین: https://escholarship.org/uc/item/19h777qd (در 28 مه 2021 قابل دسترسی است).
- Haklay، M. Citizen Science و داوطلبانه اطلاعات جغرافیایی: بررسی اجمالی و گونه شناسی مشارکت. در جمع سپاری دانش جغرافیایی ; Springer Science and Business Media LLC: Dordrecht، هلند، 2013; صص 105-122. [ Google Scholar ]
- ویگینز، ای. Crowston, K. From Conservation to Crowdsourcing: A Tipology of Citizen Science. در مجموعه مقالات چهل و چهارمین کنفرانس بین المللی هاوایی در علوم سیستمی ; موسسه مهندسین برق و الکترونیک (IEEE): Piscataway, NJ, USA, 2011; صص 1-10. [ Google Scholar ]
- وایلدرمن، سی سی شرمنز کریک: پرتره یک حوضه آبخیز. در گزارش وضعیت فنی کالج دیکینسون: کارلایل، انگلستان، 2004. [ Google Scholar ]
- براون، پ. مورلو-فروش، آر. Zavestoski, S. Contested Sillnesses : Citizens, Science, and Health Movements Social ; انتشارات دانشگاه کالیفرنیا: Berekelay، CA، USA، 2011. [ Google Scholar ]
- Shade، S. سینارکی، سی. Roglia، E. داده های علمی از و برای شهروند. دوشنبه اول 2017 22 . [ Google Scholar ] [ CrossRef ]
- کمپبل، جی. بوسر، ا. فراسل، دی. Meloche, M. Citizen Science و ادغام داده ها برای درک زباله های دریایی. در Data for Good Exchange ; IIASA: نیویورک، نیویورک، ایالات متحده آمریکا، 2019. [ Google Scholar ]
- Hsu، YC; کراس، جی. دیل، پی. تاسوتا، م. دیاس، بی. سارجنت، آر. نوربخش، آی. بوی پیتسبورگ: دانش شهروندان جامعه درگیر برای کیفیت هوا. ACM Trans. تعامل داشتن. هوشمند سیستم 2020 ، 10 ، 1-49. [ Google Scholar ] [ CrossRef ]
- کوکامن، س. Gokceoglu، C. یک برنامه CitSci برای جمع آوری داده های زمین لغزش. زمین لغزش 2018 ، 16 ، 611-615. [ Google Scholar ] [ CrossRef ]
- کوکامن، س. Gokceoglu، C. در مورد استفاده از Citsci و Vgi در ارزیابی خطرات طبیعی. ISPRS Int. قوس. فتوگرام از راه دور. حس اسپات. Inf. علمی 2018 ، XLII-5 ، 69-73. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- وودال، جنرال موتورز؛ هوور، دکتر ویلیامز، آر. بندیکت، ک. هارپر، ام. سو، جی.-سی. جرابک، ع.م. استوارت، ام جی. براون، JS; Hulla، JE; و همکاران تفسیر خوانشهای سنسور هوای موبایل و دستی در رابطه با استانداردهای کیفیت هوا و ارزشهای مرجع اثر سلامت: مقابله با چالشها. Atmosphere 2017 , 8 , 182. [ Google Scholar ] [ CrossRef ][ Green Version ]
- کن، ر. کوکامن، س. Gokceoglu، C. توسعه یک پلت فرم CitSci و هوش مصنوعی پشتیبانی شده GIS برای جمع آوری داده های زمین لغزش. ISPRS Int. قوس. فتوگرام از راه دور. حس تفت. Inf. علمی 2020 ، XLIII-B5-2 ، 43-50. [ Google Scholar ] [ CrossRef ]
- کن، ر. کوکامن، س. Gokceoglu، C. یک معماری شبکه عصبی کانولوشن برای تشخیص خودکار عکسهای زمین لغزش برای ارزیابی کیفیت دادههای اطلاعات جغرافیایی داوطلبانه و علم شهروندان. ISPRS Int. J. Geo-Inf. 2019 ، 8 ، 300. [ Google Scholar ] [ CrossRef ] [ نسخه سبز ]
- یالچین، آی. کوکامن، س. Gokceoglu، C. یک رویکرد CitSci برای نقشه برداری سریع شدت زلزله: مطالعه موردی از استانبول (ترکیه). ISPRS Int. J. Geo-Inf. 2020 ، 9 ، 266. [ Google Scholar ] [ CrossRef ] [ نسخه سبز ]
- کلیمن-فینان، ج. Scheuch، M. Winter, S. کمک های علم شهروندی به آموزش علم: بررسی یک پروژه علمی شهروندی تنوع زیستی با مدارس در اروپای مرکزی. بین المللی J. Sci. آموزش. 2018 ، 40 ، 2078–2098. [ Google Scholar ] [ CrossRef ]
- کوبوری، ح. دیکینسون، جی ال. وشیتانی، ط. ساکورای، ر. آمانو، تی. کوماتسو، ن. کیتامورا، دبلیو. تاکاگاوا، اس. کویاما، ک. اوگاوارا، تی. و همکاران علم شهروندی: رویکردی جدید برای پیشبرد اکولوژی، آموزش و حفاظت Ecol. Res. 2016 ، 31 ، 1-19. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- بانی، آر. کوپر، سی بی; دیکینسون، جی. کلینگ، اس. فیلیپس، تی. روزنبرگ، KV; Shirk, J. Citizen Science: ابزاری در حال توسعه برای گسترش دانش علم و سواد علمی. Bioscience 2009 ، 59 ، 977-984. [ Google Scholar ] [ CrossRef ]
- کومن، MH; رودریگز، ای. هافمن، آ. پترسن، سی. Oberhauser, K. علم اصیل با علم شهروندی و پروژه های نمایشگاه علمی دانش آموز محور. علمی آموزش. 2018 ، 102 ، 593-644. [ Google Scholar ] [ CrossRef ]
- بلوم، EH; Crowder، DW ترویج جمع آوری داده ها در پروژه های علمی شهروند گرده افشان. شهروند علمی نظریه Pr. 2020 ، 5 ، 5. [ Google Scholar ] [ CrossRef ] [ نسخه سبز ]
- برزسکی، KE; گونتر، ام اس; Black, JM ارزیابی جمعیت شناسی سمور رودخانه ای با استفاده از روش های ژنتیکی غیرتهاجمی. جی. وایلدل. مدیریت 2013 ، 77 ، 1523-1531. [ Google Scholar ] [ CrossRef ]
- کوکامن، س. Gokceoglu، C. CitSci به عنوان یک رویکرد جدید برای تحقیقات زمین لغزش. در مجموعه مقالات یادداشت های سخنرانی ; Springer: Cham، سوئیس، 2019; ص 161-183. [ Google Scholar ]
- داوطلبان باغ وحش Clery، D. Galaxy Pain و Glory of Research را به اشتراک می گذارند. Science 2011 ، 333 ، 173-175. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- رید، جی. رادیک، ام جی; لاردنر، آ. کارنی، ک. تحلیل عاملی اکتشافی انگیزههای مشارکت در Zooniverse، مجموعهای از پروژههای علمی شهروند مجازی. در مجموعه مقالات 2013، چهل و ششمین کنفرانس بین المللی هاوایی در علوم سیستم، Wailea، Maui، HI، ایالات متحده آمریکا، 7-10 ژانویه 2013. صص 610-619. [ Google Scholar ]
- فولت، آر. Strezov, V. An Analysis of Citizen Science Based Research: Usage and Publication Patterns. PLoS ONE 2015 , 10 , e0143687. [ Google Scholar ] [ CrossRef ] [ PubMed ][ نسخه سبز ]
- پریش، جی کی. برگس، اچ. Weltzin، JF; فورتسون، ال. ویگینز، ای. سیمونز، ب. افشای علم در علم شهروندی: تناسب با هدف و طراحی عمدی. یکپارچه سازی Comp. Biol. 2018 ، 58 ، 150-160. [ Google Scholar ] [ CrossRef ]
- برودر، LD; دیویلی، جی. ون اورز، اچ. شویت، جی. Wagemakers, A. Citizen Science برای سلامت عمومی. ارتقای سلامت. بین المللی 2016 ، 33 ، 505-514. [ Google Scholar ] [ CrossRef ]
- دانلی، ا. کرو، او. ریگان، ای. بیگلی، اس. Caffarra، A. نقش علم شهروندی در نظارت بر تنوع زیستی در ایرلند. بین المللی J. Biometeorol. 2013 ، 58 ، 1237-1249. [ Google Scholar ] [ CrossRef ]
- دامولاس، تی. هنری، اس. فارنسورث، ا. لنزون، ام. گومز، سی. بیزی طبقهبندی تماسهای پرواز با هسته تابآور زمان پویا. در مجموعه مقالات نهمین کنفرانس بین المللی یادگیری ماشین و کاربردها در سال 2010 . موسسه مهندسین برق و الکترونیک (IEEE): Piscataway, NJ, USA, 2010; ص 424-429. [ Google Scholar ]
- Stoeckle، M. Taxonomy، DNA، and the Bar Code of Life. Bioscience 2003 ، 53 ، 796-797. [ Google Scholar ] [ CrossRef ]
- ترنر، دبلیو. اسپکتور، اس. گاردینر، ن. فلادلند، ام. استرلینگ، ای. اشتاینینگر، ام. سنجش از دور برای علم و حفاظت از تنوع زیستی. Trends Ecol. تکامل. 2003 ، 18 ، 306-314. [ Google Scholar ] [ CrossRef ]
- همی، ا. گراهام، I. علم هکر در مقابل علم بسته: ایجاد زیرساخت نظارت بر محیط زیست. Inf. اشتراک. Soc. 2013 ، 17 ، 830-842. [ Google Scholar ] [ CrossRef ]
- سالیوان، پی جی؛ آچسون، جی. Angermeier، PL; فاست، تی. فلما، ج. جونز، سی ام. نادسن، EE; Minello، TJ; Secor، DH; واندرلیچ، آر. و همکاران تعریف و اجرای بهترین علوم موجود برای علوم، سیاست و مدیریت شیلات و محیط زیست. ماهیگیری 2006 ، 31 ، 460. [ Google Scholar ]
- سینگ، پی. ساران، س. کومار، دی. پادالیا، اچ. سریواستاوا، آ. کومار، نقشه برداری گونه با استفاده از رویکرد علم شهروندی از طریق پورتال IBIN: مورد استفاده در کوهپایه های هیمالیا. J. شرکت هندی از راه دور. Sens. 2018 , 46 , 1725–1737. [ Google Scholar ] [ CrossRef ]
- Novacek، MJ درگیر کردن مردم در مسائل تنوع زیستی. Proc. Natl. آکادمی علمی ایالات متحده آمریکا 2008 ، 105 ، 11571-11578. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- سیلورتاون، جی. کوک، ال. کامرون، آر. داد، م. مک کانوی، ک. ورثینگتون، جی. اسکلتون، پی. آنتون، سی. بوسدورف، او. باور، بی. و همکاران سیتیزن ساینس تغییر تکاملی غیرمنتظره در مقیاس قاره ای را در یک ارگانیسم مدل آشکار می کند. PLoS ONE 2011 ، 6 ، e18927. [ Google Scholar ] [ CrossRef ] [ PubMed ][ نسخه سبز ]
- کنراد، سی سی; Hilchey، KG مروری بر علم شهروندی و نظارت بر محیط زیست مبتنی بر جامعه: مسائل و فرصتها. محیط زیست نظارت کنید. ارزیابی کنید. 2010 ، 176 ، 273-291. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- فریمن، SN; نوبل، دی جی؛ نیوزون، SE; Baillie، SR مدل سازی تغییرات جمعیت با استفاده از داده های بررسی های مختلف: سرشماری مشترک پرندگان و بررسی پرندگان پرورشی. Bird Study 2007 ، 54 ، 61-72. [ Google Scholar ] [ CrossRef ]
- Michener، WK; جونز، MB Ecoinformatics: حمایت از اکولوژی به عنوان یک علم داده فشرده. Trends Ecol. تکامل. 2012 ، 27 ، 85-93. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- یو، جی. کلینگ، اس. گربراخت، جی. وانگ، دبلیو.-کی. تأیید خودکار داده ها در یک پروژه علمی شهروندی در مقیاس بزرگ: مطالعه موردی در مجموعه مقالات هشتمین کنفرانس بین المللی IEEE در سال 2012 در علوم الکترونیکی . موسسه مهندسین برق و الکترونیک (IEEE): Piscataway, NJ, USA, 2012; صص 1-8. [ Google Scholar ]
- McClintock، BT; بیلی، LL; پولاک، KH; Simons, TR خطای مشاهده بدون مدل باعث تعصب در هنگام استنباط الگوها و پویایی وقوع گونه ها از طریق تشخیص های شنیداری می شود. اکولوژی 2010 ، 91 ، 2446-2454. [ Google Scholar ] [ CrossRef ]
- Boakes، EH; مک گوان، PJK؛ فولر، آر. چانگ چینگ، دی. کلارک، NE; اوکانر، ک. میس، جی. دیدگاه های تحریف شده از تنوع زیستی: سوگیری مکانی و زمانی در داده های وقوع گونه ها. PLoS Biol. 2010 ، 8 ، e1000385. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- ادواردز، جی ال. لین، MA; نیلسن، ES قابلیت همکاری پایگاه های داده تنوع زیستی: اطلاعات تنوع زیستی در هر دسکتاپ. Science 2000 , 289 , 2312-2314. [ Google Scholar ] [ CrossRef ]
- Wheeler، QD چه می شود اگر GBIF؟ Bioscience 2004 , 54 , 718. [ Google Scholar ] [ CrossRef ]
- Telenius، A. اطلاعات تنوع زیستی عمومی می شود: GBIF در خدمت شماست. نورد. جی. بات. 2011 ، 29 ، 378-381. [ Google Scholar ] [ CrossRef ]
- سوارز، AV; Tsutsui، ND ارزش مجموعه های موزه برای تحقیق و جامعه. BioScience 2004 ، 54 ، 66-74. [ Google Scholar ] [ CrossRef ]
- تامل، WF; کارتر، GA; فلمونز، پ. چاپمن، RR ارزیابی داده های مجموعه موزه برای استفاده در ارزیابی تنوع زیستی. حفظ کنید. Biol. 2001 ، 15 ، 648-657. [ Google Scholar ] [ CrossRef ]
- پنیسی، ای. احیای طبقه بندی. Science 2000 , 289 , 2306-2308. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- شفر، اچ. فیشر، RN; دیویدسون، سی. نقش مجموعه های تاریخ طبیعی در ثبت گونه ها کاهش می یابد. Trends Ecol. تکامل. 1998 ، 13 ، 27-30. [ Google Scholar ] [ CrossRef ]
- Lozier, JD; آنیلو، پی. هیکرسون، امجی پیشبینی توزیع ساسکواچ در غرب آمریکای شمالی: هر چیزی با مدلسازی جایگاه اکولوژیکی مطابقت دارد. J. Biogeogr. 2009 ، 36 ، 1623-1627. [ Google Scholar ] [ CrossRef ]
- Thorn, JS; نجمان، وی. اسمیت، دی. Nekaris، KAI مدلسازی طاقچه اکولوژیکی به عنوان تکنیکی برای ارزیابی تهدیدات و تعیین اولویتهای حفاظتی برای لورهای آهسته آسیایی (Primates: Nycticebus). غواصان. توزیع کنید. 2009 ، 15 ، 289-298. [ Google Scholar ] [ CrossRef ]
- هراندو مورایرا، اس. ویتالز، دی. Nualart، N.; گومز-بلور، سی. ایبانز، ن. ماسو، اس. کاچون-فررو، پی. گونزالس-گوتیرز، PA; گیلات، دی. هررا، آی. و همکاران الگوهای توزیع جهانی و مدلسازی طاقچههای مهاجم Kalanchoe × Houghtonii (Crassulaceae). علمی جمهوری 2020 ، 10 ، 1-18. [ Google Scholar ] [ CrossRef ] [ PubMed ][ نسخه سبز ]
- پریو، سی. نادو، آی. سلیر، ی. ایسلین-نونددو، اف. برتراند، آر. کولاس، م. کاپینا، سی. مدل سازی گراندژان، اف. طاقچه برای هدایت اقدامات حفاظتی در فرانسه برای خرچنگ در معرض خطر Austropotamobius pallipes در رابطه با مهاجم Pacifastacus leniusculus. تازه Biol. 2019 ، 65 ، 304–315. [ Google Scholar ] [ CrossRef ]
- الحاجری، BH; Fourcade، Y. همبستگی بالا بین برآورد داده های محیطی در سطح گونه استخراج شده از نقشه های محدوده متخصص IUCN و از داده های وقوع GBIF. J. Biogeogr. 2019 ، 46 ، 1329–1341. [ Google Scholar ] [ CrossRef ]
- سانگ، اس. Kwon، Y.-S. لی، DK; Cho, Y. پیشبینی توزیع بالقوه گونه مهاجم، Solenopsis invicta Buren (Hymenoptera: Formicidae)، تحت تغییرات آب و هوا با استفاده از مدلهای توزیع گونهها. انتومول. Res. 2018 ، 48 ، 505-513. [ Google Scholar ] [ CrossRef ]
- Huettmann، F. آرتوخین، ی. گیلگ، او. هامفریز، جی. پیشبینیهای 27 توزیع پرنده دریایی پلاژیک قطب شمال با استفاده از متغیرهای محیطی عمومی، ارزیابی شده با دادههای مستعمره: اولین پلتفرم سنتز دسترسی باز دیجیتال IPY و GBIF. Mar. Biodivers. 2011 ، 41 ، 141-179. [ Google Scholar ] [ CrossRef ]
- گومز، VHF؛ میل، FE; گاسلینگ، WD; ویرا، ICG; Salomão، RP; Ter Steege، H. مدلسازی توزیع گونههای درختی آمازون در پاسخ به تغییرات آب و هوایی طولانیمدت در اواسط اواخر هولوسن. J. Biogeogr. 2020 ، 47 ، 1530-1540. [ Google Scholar ] [ CrossRef ]
- هاستینگز، آر. راترفورد، لس آنجلس; فریر، جی جی. کالینز، RA; سیمپسون، SD; Genner، MJ تغییرات آب و هوایی باعث افزایش قطبی و کاهش به سمت استوا در گونه های دریایی می شود. Curr. Biol. 2020 ، 30 ، 1572-1577.e2. [ Google Scholar ] [ CrossRef ] [ PubMed ][ نسخه سبز ]
- Siqueira، SDF; هیگوچی، پی. داسیلوا، توزیع جغرافیایی بالقوه معاصر و آینده Cedrela fissilis Vell. تحت سناریوهای تغییر آب و هوا Revista Árvore 2019 ، 43 ، 43. [ Google Scholar ] [ CrossRef ]
- بندر، IMA؛ بوسیدن، WD; بونینگ-گیز، ک. هنسن، آی. کوهن، آی. نواک، ال. تاپفر، تی. ویگاند، تی. Dehling، DM; Schleuning، M. اثرات پیشبینیشده تغییر آب و هوا بر تنوع عملکردی پرندگان میوهخوار در امتداد شیب ارتفاعی استوایی. علمی جمهوری 2019 ، 9 ، 1-12. [ Google Scholar ] [ CrossRef ]
- ایکدا، دی اچ. حداکثر، TL; آلن، جی. لاو، MK; شوستر، اس ام. Whitham, TG مدلهای اقلیمی بومشناختی با اطلاعات ژنتیکی پیشبینی تغییرات آب و هوا را بهبود میبخشند. گلوب. چانگ. Biol. 2017 ، 23 ، 164-176. [ Google Scholar ] [ CrossRef ] [ PubMed ]
- شارما، RK; گویال، AK; شارما، ام. اکولوژی و تکامل انگلی آشیانه در فاخته هندی. نات. محیط زیست آلودگی تکنولوژی 2015 ، 14 ، 847-853. [ Google Scholar ]
- GBIF.org. وقوع GBIF. در دسترس آنلاین: https://doi.org/10.15468/dl.gd9tk3 (در 23 آوریل 2021 قابل دسترسی است).
- فصل بادهای موسمی جنوب غربی در دسترس آنلاین: https://www.imdchennai.gov.in/swweb.htm (در 31 اکتبر 2020 قابل دسترسی است).
- بررسی بارش باران های موسمی شمال شرقی تامیلناد. در دسترس آنلاین: https://metnet.imd.gov.in/mausamdocs/1413_F.pdf (در 31 اکتبر 2020 قابل دسترسی است).
- فیک، SE; Hijmans، RJ WorldClim 2: سطوح آب و هوایی با تفکیک مکانی جدید 1 کیلومتری برای مناطق خشکی جهانی. بین المللی جی.کلیماتول. 2017 ، 37 ، 4302-4315. [ Google Scholar ] [ CrossRef ]
- داده های آب و هوا و آب و هوای جهانی در دسترس آنلاین: https://www.worldclim.org/data/index.html (در 31 اکتبر 2020 قابل دسترسی است).
- سفریان، ر. نبات، پ. میچو، م. سن مارتین، دی. ولدوایر، ا. کالین، جی. دچارم، بی. دلیر، سی. برتت، اس. شوالیه، ام. و همکاران ارزیابی مدل سیستم زمین CNRM، CNRM-ESM2-1: نقش فرآیندهای سیستم زمین در اقلیم کنونی و آینده. J. Adv. مدل. سیستم زمین 2019 ، 11 ، 4182-4227. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- آیرینگ، وی. استخوانی، اس. Meehl، GA; ارشد، کالیفرنیا؛ استیونز، بی. Stouffer، RJ; تیلور، KE مروری بر طرح آزمایشی و سازماندهی فاز 6 پروژه مقایسه مدل زوجی (CMIP6). Geosci. مدل Dev. 2016 ، 9 ، 1937-1958. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- Dormann، CF; الیت، جی. باچر، اس. بوخمن، سی. کارل، جی. کاره، جی. مارکوز، جی آر جی؛ گروبر، بی. لافورکید، بی. لیتائو، پی جی. و همکاران هم خطی: مروری بر روش های مقابله با آن و یک مطالعه شبیه سازی که عملکرد آنها را ارزیابی می کند. اکوگرافی 2013 ، 36 ، 27-46. [ Google Scholar ] [ CrossRef ]
- Naimi, B. Usdm: تجزیه و تحلیل عدم قطعیت برای مدل های توزیع گونه ها. نسخه R پکیج 1.1-18. 2017. در دسترس آنلاین: https://cran.r-project.org/web/packages/usdm/index.html (در 12 ژانویه 2021 قابل دسترسی است).
- زور، AF; Ieno، EN; Elphick، CS پروتکلی برای کاوش داده ها برای جلوگیری از مشکلات آماری رایج: کاوش داده ها. روش ها Ecol. تکامل. 2010 ، 1 ، 3-14. [ Google Scholar ] [ CrossRef ]
- Phillips, SJ آموزش مختصری در مورد Maxent. AT&T Res. 2005 ، 190 ، 231-259. [ Google Scholar ]
- Qin، A.; لیو، بی. گوا، کیو. Bussmann، RW; ما، اف. ژیان، ز. خو، جی. مدلسازی پی، اس. ماکسنت برای پیشبینی اثرات تغییرات آب و هوایی بر توزیع بالقوه Thuja sutchuenensis Franch.، یک درخت مخروطی بسیار در خطر انقراض از جنوب غربی چین. گلوب. Ecol. حفظ کنید. 2017 ، 10 ، 139-146. [ Google Scholar ] [ CrossRef ]
- دینگلیانگ، ایکس. Zhanqing، H. اصل حداکثر آنتروپی و کاربردهای آن در اکولوژی. تنوع زیستی علمی 2011 ، 19 ، 295-302. [ Google Scholar ] [ CrossRef ]
- یانگ، ایکس. کوشواها، س. ساران، س. خو، جی. مدلسازی روی، پی ماکسنت برای پیشبینی توزیع بالقوه گیاه دارویی Justicia adhatoda L. در کوهپایههای هیمالیا کوچک. Ecol. مهندس 2013 ، 51 ، 83-87. [ Google Scholar ] [ CrossRef ]
- او، جی. کولووس، A. رویکرد آنتروپی حداکثر بیزی و کاربردهای آن: بررسی. استوک. محیط زیست Res. ارزیابی ریسک 2018 ، 32 ، 859-877. [ Google Scholar ] [ CrossRef ]
- کلیدون، ا. ملحی، ی. Cox، P. حداکثر تولید آنتروپی در سیستم های زیست محیطی و اکولوژیکی. فیلوس ترانس. R. Soc. B Biol. علمی 2010 ، 365 ، 1297-1302. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- Fourcade، Y.; انگلر، جو. رودر، دی. ثاندی، جی. نقشهبرداری توزیع گونهها با MAXENT با استفاده از نمونهای با سوگیری جغرافیایی از دادههای حضور: ارزیابی عملکرد روشها برای تصحیح سوگیری نمونهبرداری. PLoS ONE 2014 ، 9 ، e97122. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- پیرسون، آر جی. Raxworthy، CJ; ناکامورا، م. پترسون، در مقاله اصلی: پیشبینی توزیع گونهها از تعداد کم سوابق وقوع: یک مورد آزمایشی با استفاده از گکوهای مرموز در ماداگاسکار. J. Biogeogr. 2006 ، 34 ، 102-117. [ Google Scholar ] [ CrossRef ]
- کومار، اس. مدلسازی Stohlgren، TJ Maxent برای پیشبینی زیستگاه مناسب درخت Canacomyrica monticola در معرض خطر و در معرض انقراض در کالدونیای جدید. جی. اکول. نات. محیط زیست 2009 ، 1 ، 094–098. [ Google Scholar ] [ CrossRef ]
- فیلدینگ، ق. Bell, JF مروری بر روشهای ارزیابی خطاهای پیشبینی در مدلهای حضور/غیاب حفاظتی. محیط زیست حفظ کنید. 1997 ، 24 ، 38-49. [ Google Scholar ] [ CrossRef ]
- Swets, JA اندازه گیری دقت سیستم های تشخیصی. علوم 1988 ، 240 ، 1285-1293. [ Google Scholar ] [ CrossRef ][ نسخه سبز ]
- گوهاتاکورتا، پ. راجیوان، م. سیکا، DR; Tyagi، A. تغییراتی را در بارش باران های موسمی جنوب غربی بر فراز هند طی سال های 1901-2011 مشاهده کرد. بین المللی جی.کلیماتول. 2015 ، 35 ، 1881-1898. [ Google Scholar ] [ CrossRef ]
- چییب، ع. بادو، وی. بوئه، جی. چویین، آی. دلیر، سی. دوفرنه، ای. فرانسوا، سی. گریتی، ES; لگی، م. پیج، سی. و همکاران تأثیرات تغییر آب و هوا بر محدوده درختان: مقایسه مدلها درک و تعیین کمیت عدم قطعیت را تسهیل میکند. Ecol. Lett. 2012 ، 15 ، 533-544. [ Google Scholar ] [ CrossRef ]

شکل 1. تصویر Clamator jacobinus (فاخته ژاکوبین یا فاخته Pied) (منبع: Pied Cuckoo Macaulay Library ML32455551).
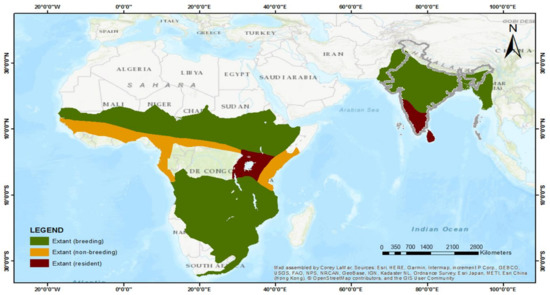
شکل 2. نقشه موجود Clamator jacobinus (Pied cuckoo) (منبع تصویر: مرجع [ 30 ]).
شکل 3. پیش بینی مناسب بودن زیستگاه در ماه های ژوئن تا سپتامبر برای محدوده فاخته Pied.
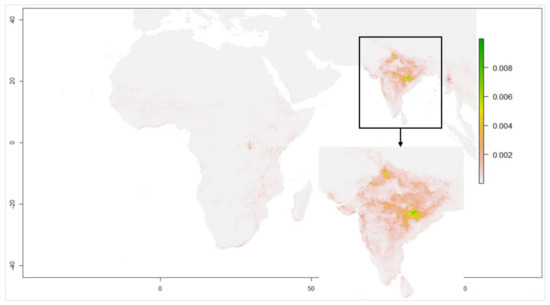
شکل 4. مدل سازی مناسب زیستگاه در سال های 1991-2005 برای دوره ژوئن-سپتامبر.
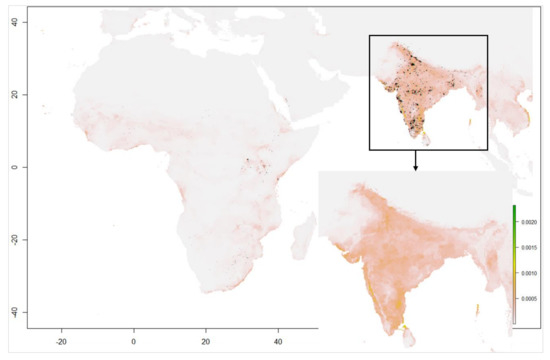
شکل 5. مدل سازی مناسب زیستگاه در 2006-2020 برای دوره ژوئن-سپتامبر.
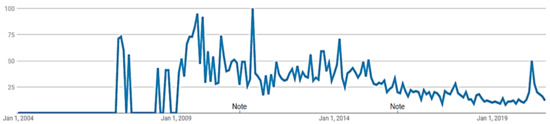
شکل 6. گرایش های گوگل از «انبوه سپاری» بر اساس جستجو در هند.

شکل 7. پیش بینی های مناسب زیستگاه فعلی در ماه های اکتبر-دسامبر برای محدوده فاخته ژاکوبین. مناطقی که با علامت ضربدر در قسمت پایین سمت راست شکل مشخص شده اند عبارتند از: آندرا پرادش (علامت آبی)، عمدتاً در مناطق ساحلی، کرالا (نقطه سیاه)، پودوچری (علامت قرمز)، رایالاسیما (علامت سبز)، کارناتاکا جنوبی. (نشان مارون) و تامیل نادو (نشان صورتی).
شکل 8. پیش بینی تناسب زیستگاه فعلی ژانویه تا مه برای محدوده فاخته Pied.
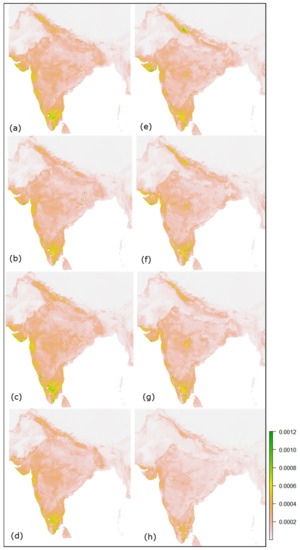
شکل 9. نقشههای مناسب زیستگاهی آینده فاخته ژاکوبین در طول ژوئن تا سپتامبر هند برای SSPs 2.6، 4.5، 7.0 و 8.5 ( a – d ) 2030 و 2050 SSPs 2.6، 4.5، 7.5.0 و 8 .
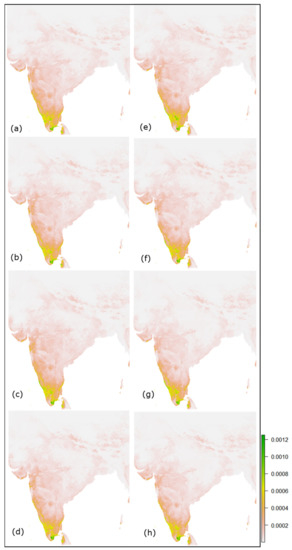
شکل 10. نقشه های مناسب بودن آینده برای فاخته های ژاکوبین از اکتبر تا دسامبر: ( a – d ) و ( e – h ) به ترتیب نشان دهنده SSPs 2.6، 4.5، 7.0 و 8.5 برای سال های 2030 و 2050 است.
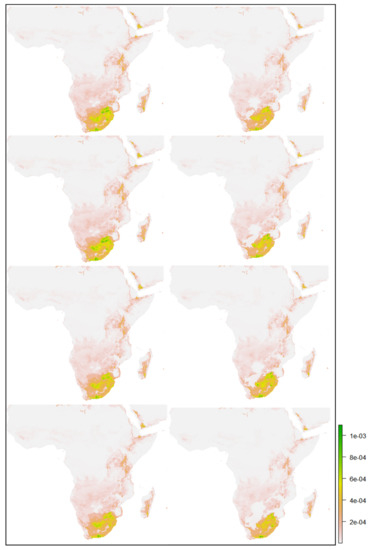
شکل 11. نقشه های آینده پیش بینی شده مناسب بودن فاخته های ژاکوبین از اکتبر تا دسامبر در آفریقا: ( a – d ) و ( e – h ) به ترتیب نشان دهنده SSPs 2.6، 4.5، 7.0 و 8.5 برای سال های 2030 و 2050 است.
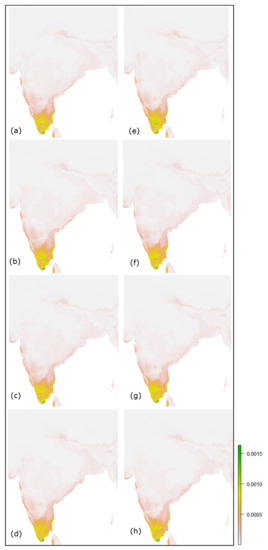
شکل 12. نقشه های تناسب آینده پیش بینی شده برای ژانویه-مه در هند: ( a – d ) و ( e – h ) به ترتیب نشان دهنده SSPs 2.6، 4.5، 7.0 و 8.5 برای سال های 2030 و 2050 است.
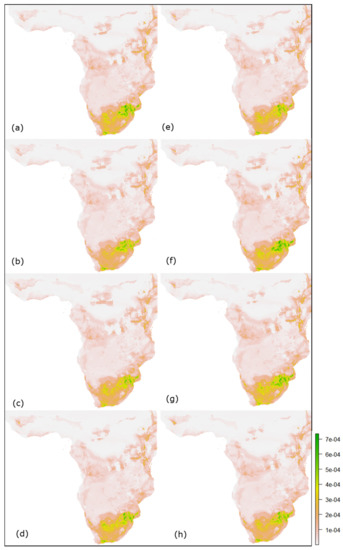
شکل 13. نقشه های مناسب آتی پیش بینی شده برای ژانویه-مه در آفریقا: ( a – d ) و ( e – h ) به ترتیب نشان دهنده SSPs 2.6، 4.5، 7.0 و 8.5 برای سال های 2030 و 2050 است.


بدون دیدگاه